8. Digestion
Chapter 8. Digestion
Kim Cross
St. Peter’s College
Muenster, Saskatchewan
8.1 Introduction
8.2 Ingestion
8.3 Motility and Propulsion
8.4 Digestion
8.5 Absorption and Assimilation
8.6 Elimination of Wastes
8.7 Summary
8.8 Suggested Readings
8.9 Glossary
8.1 Introduction
One of the main principles of life is that you need energy to maintain the organised structures of your cells and body. You also need raw materials for growth and repair. For plants, this seems easy: sit in the sun, and absorb carbon dioxide, water, and a few raw minerals to make the energy-rich, complex, large molecules that are the building blocks of all living things. However, animals get most of their energy and nutrients from pre-existing sources that are living or were once living. In other words, animals are heterotrophic (Greek, héteros: “other”; trophé: “nourishment”). So it seems logical that animals would develop a system specialized for taking up chemical energy and nutrients: this is the digestive system.
Let’s briefly explore the question, what is a nutrient?
A nutrient can sometimes be difficult to define, since nutrients can be classified differently depending on their usage and importance, their source, and their chemical properties. Considering their usage and importance, a nutrient is any substance needed to maintain homeostasis and metabolic activities necessary for life. Apply this to your own body; what do you need to maintain your body over the course of a day?
Water, carbon, nitrogen, phosphorous, sulphur, sodium, potassium, iron, magnesium, zinc, glucose, protein, adenine, cholesterol, ascorbic acid, and niacin.
You have probably realized that you need these and many more nutrients to maintain homeostasis and metabolism.
The general source of nutrients is an animal’s food, or diet. Over the course of an animal’s life, the diet may be solid foods, liquid foods, or a combination of both. The chemical properties of the diet will vary from food source to food source. Some nutrients in the diet are considered inorganic, others are considered organic.
We’ve been using the words “organic” and “inorganic” without defining them. What is the difference between an organic substance and an inorganic substance? There isn’t a precise definition, but generally, most molecules that contain the element carbon are considered organic. The exceptions are the very simple compounds where the carbon is fully oxidised, such as the gas carbon dioxide. These, along with molecules that don’t contain carbon, are considered “inorganic”.
An animal’s diet will contain both essential and non-essential nutrients. Understand, all nutrients are important, and saying a nutrient is essential doesn’t imply that it is more important than a non-essential nutrient. Instead, an essential nutrient is a nutrient that an animal cannot make through its own metabolism. The animal must get the nutrient from its diet. By contrast, an organism can make enough of each of the non-essential nutrients during normal metabolism. Most inorganic nutrients are essential. Can you make an iron atom in your body? However, only some organic nutrients are essential.
Many of the essential organic nutrients are called vitamins. Others are certain amino acids and certain fats that must be obtained in the diet. No one group of animals requires the same essential nutrients. For example, it is well documented that humans and many frugivores (fruit eaters) require ascorbic acid – vitamin C – while most other animals can make their own ascorbic acid. See Table 8.1 for a brief list of essential nutrients and their function within the human body.
Table 8.1 Major essential nutrients required in the human diet, their dietary sources, and their functions in the body.
| Nutrient | Source | Function |
| Water | Diet and metabolism | Solution for metabolism and transport, 70% of body mass. |
| Nine amino acids: phenylalanine, valine, threonine, tryptophan, methionine, leucine, isoleucine, lysine, histidine | Meat, dairy, legumes (peas and beans), nuts, general diet | Protein production. Brain development of young children is particularly affected by protein deficiency. |
| Calcium | Dairy, grains, nuts, fish with bones, tofu | Bone formation, intracellular signalling especially in the nervous system and muscle. |
| Phosphorus | Meat, dairy, grains, nuts | Bone. Nucleic acids, ATP, cell membranes. |
| Potassium | Fruit, nuts, leafy green vegetables, dairy, meat | Cation inside cells. Important in nerve and muscle. Especially osmoregulation and heart rhythm. |
| Sodium | Table salt, meat, dairy | Cation in fluids outside cell. Controls blood volume, osmoregulation, secondary active transport in the gut. |
| Iodine | Table salt, seafood, plants from iodine-rich soil. | Thyroid hormone production, which regulates metabolic rate. |
| Iron | Meat, leafy green vegetables, peas and beans | Respiratory gas transport – haemoglobin, myoglobin. Cellular respiration. |
| Zinc | Meat, nuts, peas and beans | Immune system function, growth, sensory function |
| Thirteen vitamins | Leafy green vegetables, meat, dairy, fruit, grains, gut bacteria | Needed in trace amounts for a wide range of metabolic processes |
| Two fatty acids: alpha-linolenic (omega-3), linoleic (omega-6) | Fish oils and other seafood, seed oils | Skin, immune system, brain, and heart function. |
Being heterotrophic implies animals must consume foods to obtain energy. Remember how plants store energy from the sun? Plants store energy by making the gas, oxygen, which they release into the atmosphere, at the same time making organic chemical compounds such as sugar, starch, and oil. It is by combining oxygen with chemicals that contain reduced carbon and hydrogen that living things are able to release this stored energy. The chapter on respiration covers how animals get the oxygen. To get the organic chemicals, most animals are completely dependent on eating.
Evolution has led to many methods of consumption, and the exact niche an animal fills in an ecosystem varies enormously between different animal species. As a result, people often define animals based on their main method of consumption. If animals obtain energy and nutrients directly from photosynthetic (or chemosynthetic) organisms, these animals are primary consumers. If the animal eats plants, it is called a herbivore. There are many different specialized kinds of herbivore. For example, frugivores focus on eating fruits, while granivores focus on grains (or seeds). However, not all primary consumers live entirely by eating: Some primary consumers are symbiotic (Defined in Chapter 1), supporting the photo- or chemo-synthetic organism within the animal’s body.
Other animals don’t get their energy and nutrients directly from primary producers. Those animals are secondary consumers, tertiary consumers, etc. If an animal obtains energy and nutrients by eating other animals, it is called a carnivore. Some carnivores are predators, killing other animals to eat, while other carnivores are scavengers, eating animals that are already dead. Some animals are parasites, feeding on other living animals without killing them. Many animals are detritivores, focusing on eating rotting plant matter, animal remains, or fecal matter. Other animals have a varied diet, consuming food from many of the previously listed sources, and these animals are called omnivores.
Not all animals eat solid food. Suspension feeders (also known as filter feeders) strain small particles or organisms from the water. Liquid feeders consume fluids, for example, blood, nectar, or sap.
If consumption methods are diverse, digestive systems must also be diverse. The most common digestive system is a tubular gut with two separate openings, the mouth and the anus. This is the “pipe” body plan mentioned in Chapter 1. The tubular gut is also called a gastrointestinal system or alimentary canal. In most larger animals, the gut is surrounded by accessory organs also considered to be part of the digestive sysem(salivary glands, liver, gall bladder, pancreas, and a few others).
Other animals, like cnidarians (sea anemones and jellyfish) and flatworms have a digestive cavity with only one opening, the mouth. A system with only a mouth and no anus is called a gastrovascular cavity. The gastrovascular cavity is often not only dedicated to digestion, but used for many other purposes such support and circulation. Sponges lack a digestive system: Their cells ingest food directly from the water they pump through the sponge’s body, and digestion occurs within the cells (intracellular digestion, see Section 8.4 – Digestion).
The previous discussion should give you an appreciation of the diversity of digestive systems and consumption methods in the animal kingdom. Yet what is consumption? Is it simply digestion? Is it just putting food in a mouth? No, consumption is a series of processes including ingestion, chewing, mixing, propulsion, chemical digestion, absorption and assimilation, and elimination of wastes (defecation). Considering all these processes, maybe we should rename the digestive system, the consumption system! The remainder of this chapter will explore all the consumption processes and how the different parts of the digestive system communicate through signals to control these processes. To simplify this discussion, the main model in this chapter will be an omnivorous mammal, the human, with its gastrointestinal tract and accessory organs. Figure 8.1 is an overview of the human digestive system. As the discussion continues throughout this chapter, please refer to this figure if you are unfamiliar with the general anatomy of a digestive system. While the human system will be our focus, we will discuss digestive systems of other animal species from time to time.
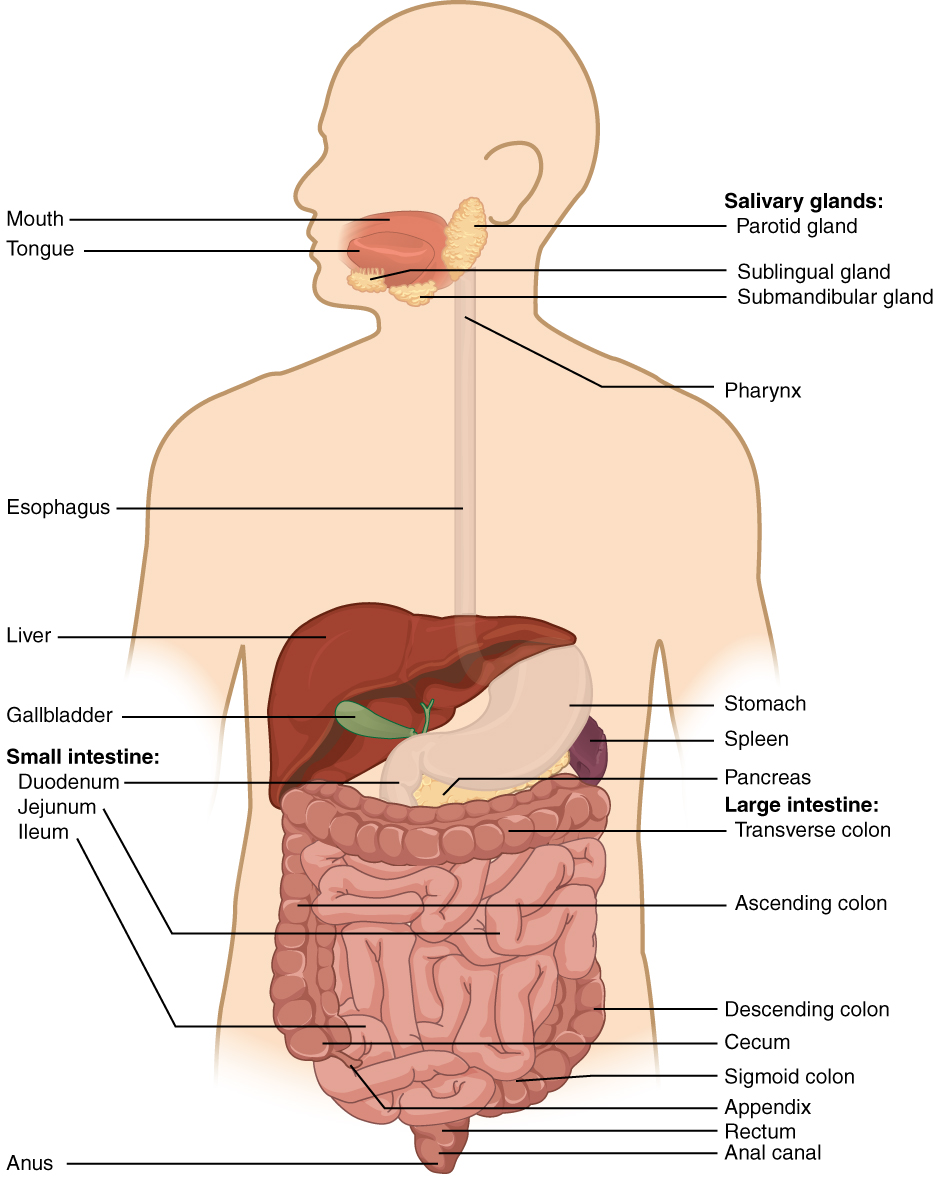
Figure 8.1 Anatomy of the human gastrointestinal tract. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
8.2 Ingestion
Ingestion, the first step, is the act of taking food into the digestive system. This typically occurs at the mouth, or oral cavity. Due to the diversity of food sources, many different structures have evolved in and around the mouth to help ingestion. The mammalian set of teeth (their dentition, Figure 8.2) includes incisors and canines for slicing and piercing. Whales have replaced their dentition with keratinized plates, called baleen, to filter feed. Elephants use an extension of their nose, the trunk, to pick up food and bring it to the mouth. The mouth of most cnidarians is surrounded by tentacles with stinging cells to immobilize their prey. Sponges lack a single mouth, instead they use flagella to draw water through many pores in their bodies. Often scientists can determine the preferred food of an animal simply by studying structures for ingestion. For example, is a mammal that lacks canines more likely to be a herbivore or a carnivore? Will an insect with long piercing mouth parts be a fluid feeder or a suspension feeder?
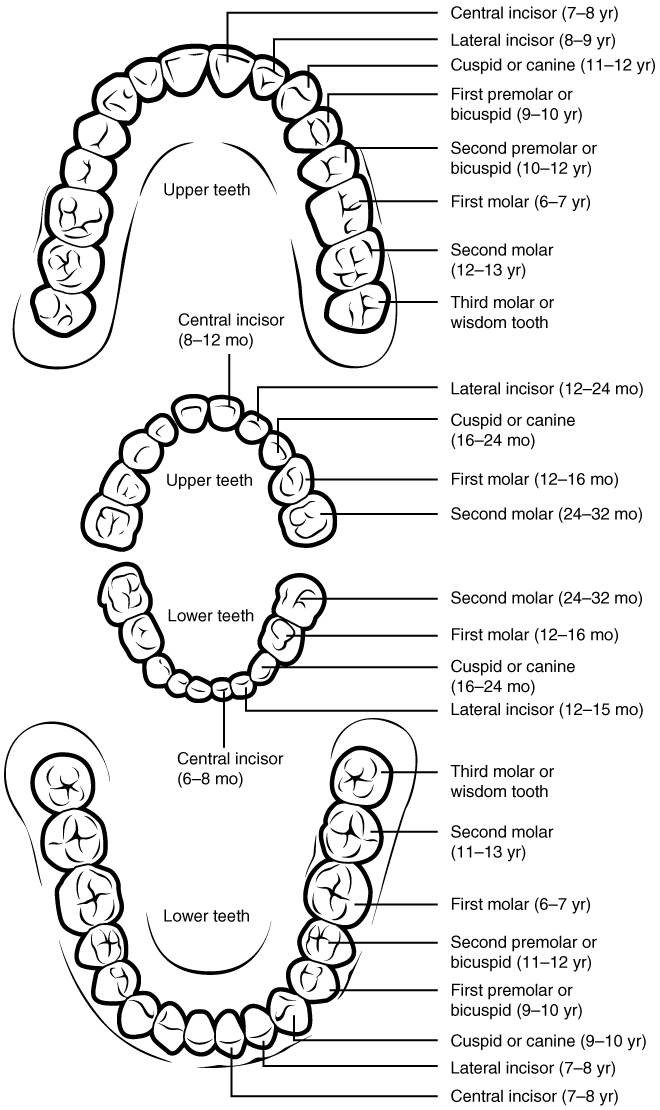
Figure 8.2 Can you guess which animal these teeth belong to? Human dentition. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
In most vertebrates, the tongue assists in ingestion, propulsion (see Section 8.3), and digestion (see Section 8.4) of food. How is the tongue involved with ingestion? Hummingbirds drink nectar using their very long, complicated tongue as a highly specialised fluid pump. Many frogs and lizards have long sticky tongues or prehensile tongues to catch insects, and many herbivores wrap their tongue around leaves and twigs to pull them free of the plant. The tongue is very sensitive; containing chemoreceptors to detect taste, proprioceptors to detect position, other mechanoreceptors to detect touch, and nociceptors to detect damaging conditions such as extreme heat or cold. Taste is the ability to detect certain chemicals in the mouth. Chemoreceptors are required for the sensation of taste, and are localized into regions on the tongue known as taste buds. There are five primary taste stimuli: sweet, salty, sour, bitter, and umami. Any stimulus from taste buds is sent via the facial cranial nerves to the thalamus, and then on to the gustation centers within the cerebral cortex. The facial nerves only receive sensory information from the anterior 2/3 of the tongue (see Figure 8.4). Any guesses as to the location of most taste buds on your tongue? The heightened sensitivity of the tongue allows animals to judge the quality and safety of the ingested food. When you put food in your mouth that is too hot, or food that has spoiled, what is your response?
8.3 Motility and Propulsion
If you ever have the chance to visit Philadelphia, stop by the Mütter Museum, where among other things, you can find slices of Albert Einstein’s brain (perfectly normal, in case you’re wondering) and a whole mounted skeleton of the unfortunate Harry Eastlack, whose muscles were gradually replaced by bone (perfectly abnormal, like something from a science fiction movie). But what is this strange object, which looks like a monstrously large sausage, 24 cm across, and almost 3 metres long?

Source: The College of Physicians of Philadelphia Digital Library, accessed April 10, 2017, http://www.cppdigitallibrary.org/items/show/4378. CC BY-NC-SA 3.0.
It’s a reminder of the importance of gut motility, that’s what it is. It’s a colon where the enteric nervous system was missing for part of its length. The individual already had a noticeably bulging belly as a baby, and his abdomen kept on growing until he toured as “Balloon Man” as a young adult. The section of colon that lacked nerve cells couldn’t contract to move material along, so by the time he died of constipation, 18 kg of feces had accumulated in his colon.
As ingestion continues, the food must be moved to locations where digestion can occur. Motility and propulsion is the movement of food through the digestive system. Many different muscle groups are responsible for motility and propulsion. The muscles within the digestive system are mostly smooth muscle under involuntary nervous control, but regions of voluntary skeletal muscle do exist. Lips, tongue and the external anal sphincter are largely comprised of voluntary muscles. Most of the internal tubular regions of the digestive system, the oesophagus, stomach, and intestines, have layers of smooth muscle groups along their entire length. Within the tube walls, smooth muscle fibre orientation may be circular, longitudinal, and oblique. This mixture of muscle orientations contributes to efficient propulsion of food via rhythmic muscle contractions.
Muscles generate the necessary force to move food in the digestive system. However, the diet often consists of dry food, and muscle action alone will not move dry food through the system. Dry food requires lubrication to move through the digestive system, otherwise blockages will occur. Lubrication is provided by mucus from a series of accessory glands and mucus secreting cells found throughout the entire length of the alimentary canal.
Our first introduction to digestive accessory organs and glands will be the salivary glands. Figure 8.3 shows the location of three main salivary glands in the human mouth; the sublingual (under the tongue), the submandibular (under the mandible or jaw) and the parotid (between the jaw and the ear). All three secrete saliva into the mouth. Next time you are in front of a mirror, lift your tongue to the roof of your mouth, and you should observe two small tubes (salivary ducts) that empty about half way between the lower incisors and the base of the tongue. These ducts lead from the submandibular salivary gland. Recall, glands using ducts to secrete materials are called exocrine glands.
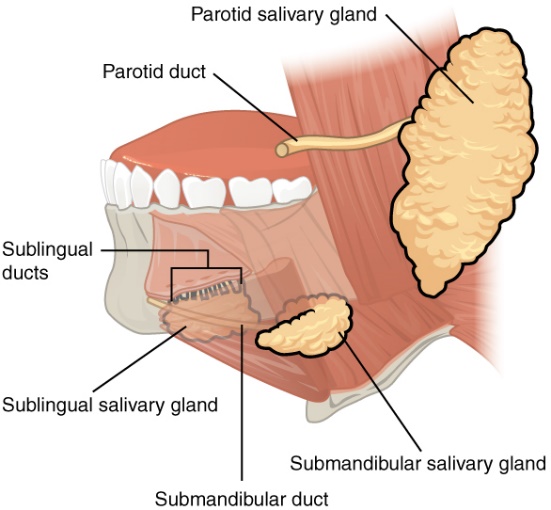
Figure 8.3 Human salivary gland position around the mouth. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
The composition of saliva can change over time, but it is mostly water and electrolytes, with a mixture of enzymes and antibacterial compounds. The main enzyme present is amylase; it begins carbohydrate digestion in the mouth (see Section 8.4). Since the mouth is an excellent environment for bacterial growth (warm, moist and lots of nutrients), antibacterial compounds reduce bacterial growth and help prevent tooth decay. By mixing the food with saliva, food is more easily digested and more easily manipulated. Once combined with saliva the food can be formed into a small, lubricated ball called a bolus (Greek: bôlos, lump), which is more easily swallowed.
Salivary glands involuntarily produce saliva throughout the day. This continuous saliva production is called resting saliva, and it may be stimulated by tongue movements, jaw movements, talking and swallowing of saliva. These stimuli travel via cranial nerves to salivary centers in the brain stem, and possibly further to the hypothalamus. In response to these stimuli, the parasympathetic division of the peripheral nervous system (PNS) releases acetylcholine via cranial nerves, exciting the salivary glands.
Upon ingestion of food, the presence of food in the mouth provides more mechanical and chemical stimuli. Increased stimulation will generate reflex arcs that bypass the salivary centres and greatly increase the flow of saliva from the resting state. Quick test: what cranial nerve is responsible for triggering saliva production in the sublingual and submandibular glands, if the stimulus that triggers the saliva production is chemical? Does it make sense that this would be the facial nerve, since it is the same nerve carrying taste information from the tongue?
Additionally, an increase in saliva production can be a conditioned reflex. Have you heard of Pavlov’s experiments on dogs? These experiments demonstrate a conditioned, feed-forward response. Organisms can be conditioned to respond quickly to certain stimuli, or certain nervous pathways can be reinforced or amplified to make processes more efficient in response to certain stimuli. When you see delicious food or smell it, your brain is conditioned for these stimuli and will anticipate that saliva is needed. As a result, “your mouth waters” at the sight or smell of food. Preparing the mouth for food is more efficient than waiting for the food to be present, and then reacting.
The production of saliva is also controlled via the sympathetic division of the PNS. Unlike other regions of the body, the sympathetic is not completely antagonistic to the parasympathetic division for saliva production. However, when salivary glands receive norepinephrine from the sympathetic pathways the saliva contains less water and is produced in lower volume. This occurs when food texture or chemistry changes, or when the animal experiences more stress. Have you experienced a “dry mouth” when writing an exam, or when exercising? Note: the production of saliva is variable in many species, and the parasympathetic and sympathetic controls may act differently from animal to animal.
Food must travel efficiently throughout the entire digestive tract. Therefore, lubrication does not stop with saliva production in the mouth. As mentioned above, the oesophagus is lined with mucous glands that release mucus to lubricate the bolus after swallowing. The stomach and intestines have mucous glands and goblet cells (epithelial cells which line the gut and release mucus), to continue lubrication in the lower portions of the digestive tract. Production of these secretions are largely under the control of the parasympathetic nervous system, but can be influenced by local events. In the stomach, the bolus is broken up and eventually becomes a soupy mixture known as chyme. This will be discussed more in Section 4 – Digestion, so let’s return the discussion to the muscles involved in motility.
Within in the oral cavity or mouth, the tongue’s voluntarily movement is started when mechanical stimuli (pressure, stretch and touch) travel through the trigeminal cranial nerve and other cranial nerves to the brain stem (medulla, pons, reticular formation). The brain stem sends impulses through motor neurons, mostly within the hypoglossal cranial nerves, to stimulate movement of the lingual (tongue) muscles. As the tongue moves it mixes food with saliva, presses the food against the teeth, and is responsible for forming the bolus.
After food is partially digested and lubricated within the mouth, it must be swallowed. The initiation of swallowing can be both voluntary and involuntary, but once swallowing begins it is a reflex. The reflex pathway begins with the tongue creating and pushing the bolus to the back of the mouth. This region is known as the pharynx region, and is often a shared pathway with the respiratory system. Within the pharynx region are many pressure receptors and stretch receptors, which send information through cranial nerves, like the glossopharyngeal and vagus nerves. The swallowing reflex is transmitted back through the hypoglossal and vagus nerves resulting in motor activity within the muscles of the tongue, pharynx, larynx (voice box) and oesophagus. Figure 8.4 is a summary of tongue innervation by cranial nerves.
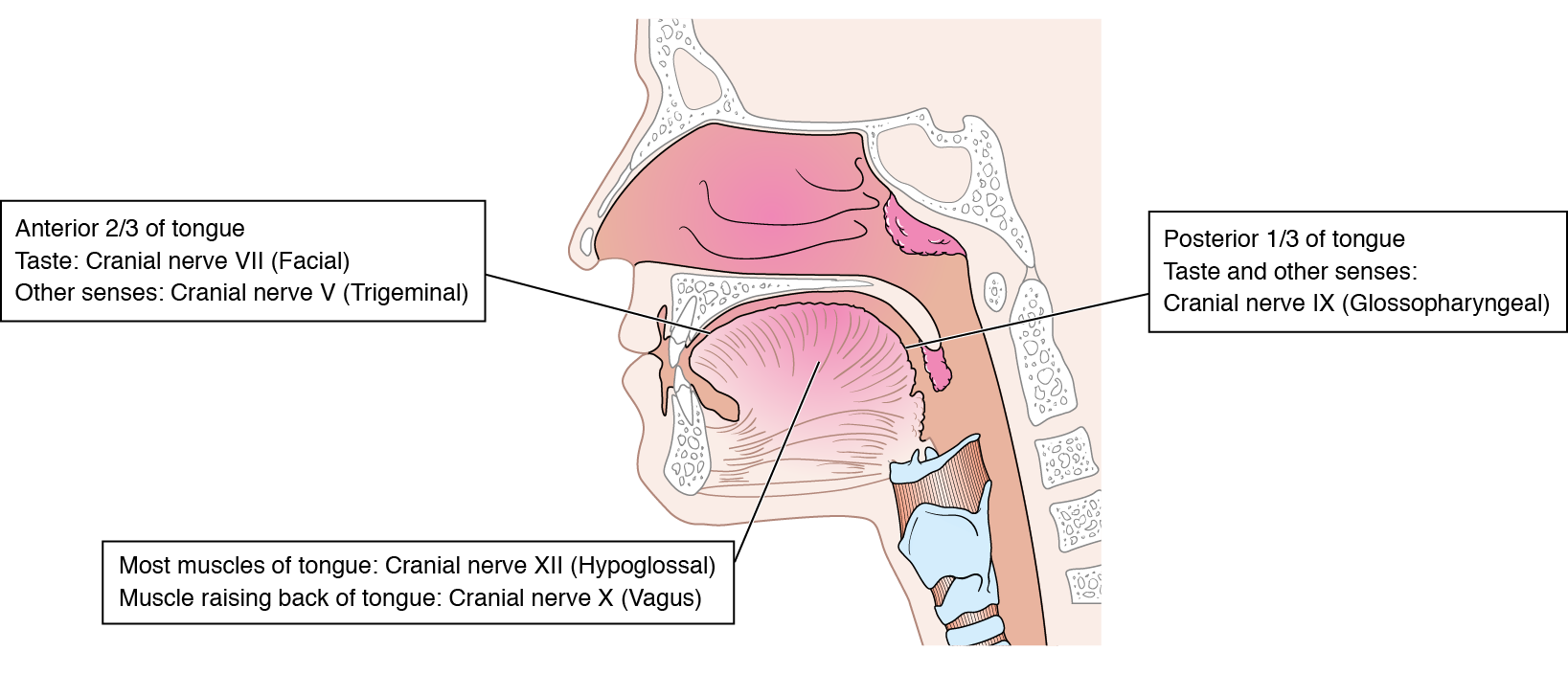
Figure 8.4 Cranial nerve innervation of the human tongue. Source of drawing: Cancer Research UK / Wikimedia Commons CC BY-SA 4.0, 2014.
Many people think the act of swallowing stops once the bolus is in the oesophagus; actually, swallowing continues to move the bolus through the oesophagus to the stomach. Within the oesophagus, the bolus causes stretching, and stretch receptors continually send information to the swallowing centers in the brain stem. The vagus nerve relays information back and forth from the oesophagus to the brain stem, ultimately resulting in a wave of muscle contractions occurring behind the bolus. These wave-like muscle contractions, known as peristalsis, do not stop until the bolus has reached the stomach. Notice this is very similar to an action potential within a nerve, once peristalsis starts it cannot be stopped and the wave is unidirectional, from the oesophageal aperture (opening) to the stomach. Figure 5 shows peristaltic contraction within the oesophagus.
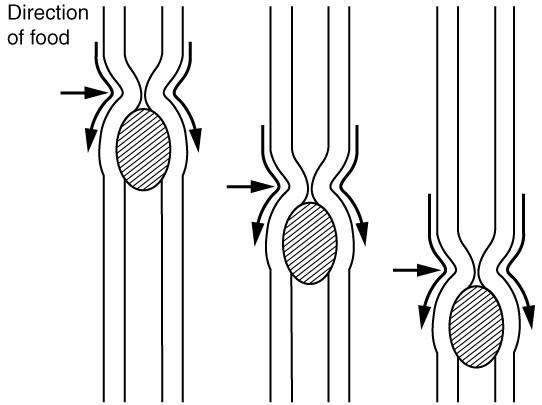
Figure 8.5 Oesophageal peristalsis causing the unidirectional movement of the bolus. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
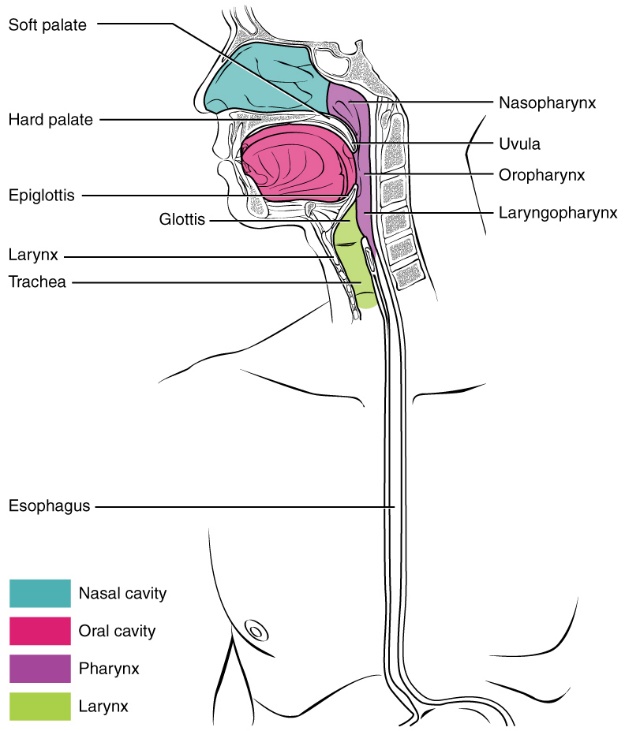
Figure 8.6 Human pharyngeal region and location of the glottis in relation to the oesophageal aperture. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
Sometimes, the direction of peristalsis in the stomach and oesophagus can be reversed. Can you think of examples when this happens? Vomiting is a common reaction when the nervous system detects an excessive amount of a toxic chemical, such as alcohol. Regurgitation is also used by many birds and some mammals as a way to feed their young.
There are two common problems with swallowing. The first concerns the pharynx and its dual role. The airway leading from the nose through to the larynx and trachea intersects the digestive system near the oesophageal aperture (opening to the oesophagus). Looking at Figure 8.6, you will notice the entrance to the airway, called the glottis, is positioned in front of the oesophageal aperture. During swallowing this means that food first encounters the airway before travelling to oesophagus. The glottis must be covered to prevent blockage of the airway, or choking. During the swallowing reflex, a small flap of cartilage, called the epiglottis, covers the glottis. The closure is accomplished through a combination of pressure of the bolus on the epiglottis, and the elevation of the larynx into the nasopharynx region. The second concern is dry, unlubricated food lodging in the oesophagus. If this occurs, excessive stretching starts a localized reflex arc. This localized reflex does not involve the CNS or PNS; rather it is mediated by two local nerve nets called the intrinsic nervous plexuses. Collectively these plexuses are called the enteric nervous system (ENS). Stimulation from the ENS results in extra mucus secretion and even stronger peristalsis. With increased lubrication and strong muscle action the stuck bolus will eventually dislodge. View the following online video for a summary of all activities during swallowing:
Swallowing pineapple juice. Source: Martin Uecker, Biomedizinische NMR Forschungs GmbH, http://www.biomednmr.mpg.de CC BY-SA 3.0
Muscle action and propulsion throughout the rest of the digestive system continues to be stimulated by the parasympathetic (acetylcholine release) and inhibited by the sympathetic (norepinephrine release) nervous system. The ENS extends throughout the digestive system and can control activity in the lower digestive system as well. For instance, if too much food is being passed from the stomach to the small intestine, stretch receptors in the small intestine will trigger a reflex that stops peristalsis in the stomach. This will give the small intestine more time to digest the food, rather than simply pushing the food through the system. This same message is reinforced by hormones like cholecystokinin (CCK) and secretin (see below). Constant stretching of the lower digestive system will eventually be relayed to the CNS, via the vagus nerve and through hormones, like CCK. As these messages increase in intensity, satiety centers in the hypothalamus will be eventually be stimulated and the feeling of being full (satiation) will bring an end to ingestion and swallowing. There may be a delay of up to 30 minutes before the satiety centers are stimulated. Therefore, eating too much food too quickly can lead to distension, or over-stretching of the stomach and intestines. Have you ever experienced the pain of a distended stomach due to over-eating during a festive period?
It is important to note that the pattern of propulsion after the oesophagus and stomach changes. Peristalsis can still occur, but it is more often replaced by segmentation in the small intestine, and haustral contractions in the large intestine. These contractions are not peristaltic. See Figure 8.7. Recall, peristalsis is a single contraction that moves like a wave, pushing the bolus from the beginning of the oesophagus to the end of the oesophagus (contrast Figure 8.7 with Figure 8.5). Segmentation and haustral contractions are simultaneous contractions, which occur at many different locations at the same time, resulting in the formation of regular pockets of nutrients and wastes along the entire intestine. Why do you think this is important in these regions? Comparing Figure 8.5, peristalsis to Figure 8.7, segmentation, you will notice that during segmentation food/chyme can move both forward and backward within the intestines, i.e. it is continually mixed. The chyme will combine more readily with digestive juices, promoting digestion. The chyme also spends more time within the intestine, promoting absorption of nutrients. In the large intestine, the pockets (haustra) are larger and this process is even slower. This likely allows bacteria to finalize digestion. Some animals can even reverse peristalsis to hold waste in the large intestine for longer, allowing bacteria even more time to finish digestion. Current research is showing the importance of the microflora in our guts, and gut microflora will be briefly discussed in the next section.
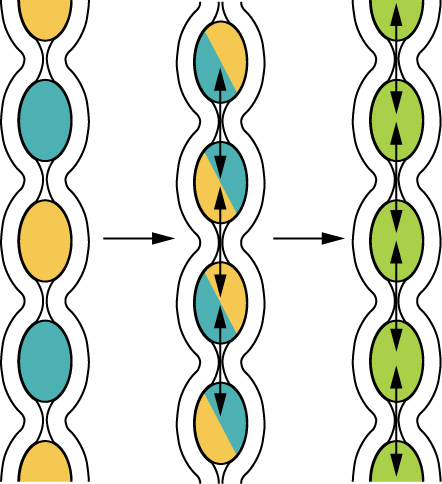
Figure 8.7 Segmentation in the small intestine. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
The final propulsive force within the digestive system, the defecation reflex, will be discussed in Section 8.6.
Throughout the digestive system, there are several junctions where localized muscles create restriction points called valves or sphincters. These restriction points control the flow of materials from one region to the next (Table 2).
Table 8.2 Restriction points in the human digestive tract, and their nervous system control
| Restriction Point | Function | Regulation |
| Lips | Initial ingestion | Voluntary |
| Oesophageal aperture | Opening to the oesophagus involved in swallowing | Involuntary |
| Cardiac sphincter | Regulates flow between the oesophagus and stomach, preventing backflow from the stomach | Involuntary |
| Pyloric sphincter | Controls flow of chyme between the stomach and duodenum (first part of the small intestine) | Involuntary |
| Ileocecal valve | Controls flow between the small and large intestine, preventing backflow from large intestine | Involuntary |
| Rectal sphincter or internal anal sphincter | Allows feces to move from rectum to anal cavity | Involuntary |
| External anal sphincter or anus | Allows feces to leave the body | Voluntary |
Finally, propulsion throughout the digestive system is not just under the direct control of the nervous system. Many hormones play important roles. Three small peptide hormones, targeting G-protein coupled receptors in muscle, play large roles in the motility of the digestive system:
- Gastrin – stimulated by presence of HCl and protein within the stomach, inhibited by extremely low pH in the stomach. Gastrin released by the stomach stimulates muscles of the stomach and the lower intestinal system.
- Secretin – stimulated by an increase in acid conditions and protein digestion within the duodenum, inhibited by return to neutral pH. Secretin released by the duodenum inhibits muscles of the stomach.
- Cholecystokinin (CCK) – stimulated by presence of fats and proteins in the duodenum and by parasympathetic division (vagus nerve and acetylcholine), inhibited by somatostatin and pancreatic peptides. CCK released by the duodenum inhibits muscles of the stomach.
8.4 Digestion
After ingestion, food must be digested to release the nutrients. Digestion is the act of breaking down food. Digestion occurs both mechanically and chemically. What is the difference between these mechanisms, and why are they both required within a gastrointestinal system?
How are the mechanisms different?
Mechanical digestion is any physical process that breaks large pieces of food into smaller pieces of food. These actions may include grinding, slicing and general mixing.
Chemical digestion is any physical process that acts primarily on the chemical bonds in the food molecules. Both mechanical digestion and chemical digestion are important. Mechanical digestion is usually needed to increase the surface area of food exposed to water and enzymes, and make sure that the food particles are small enough to be thoroughly mixed with the digestive juices. Chemical digestion is needed to make the food molecules small and simple enough to dissolve in water and be absorbed by the cells of the small intestine.
Figure 8.8 Only pythons swallow their food whole. If you chew your food, the volume stays the same, but the surface area increases. Source: Mothers for Mastication of Meals, https://mmm.chewyourfood.org CC BY 3.0, 1917.
Why are both mechanical and chemical digestion required? By now, you have likely noticed a common theme through many biology courses and textbooks; it is the concept of surface area to volume ratio. Any time the surface area of an object can be increased more activity can occur on the greater surface.
It should be clear that many of the same structures used for ingestion, motility and propulsion also assist with mechanical digestion. Mechanical digestion begins in the mouth with chewing (mastication), and requires the action of the tongue and teeth. Recall from earlier discussions that the tongue contributes to mechanical digestion by moving the food around the mouth and mixing saliva with the food. The tongue also keeps food pressed against teeth increasing mechanical digestion. The chewing dentition is typically found toward the back of the mouth, consisting of premolars and molars. Again, scientists can often determine the types of foods consumed by animal by studying the structure of the pre-molars and molars. Large molars with flat surfaces are useful for grinding tough plant fibres. Molars with high, sharp cusps act like scissors and are useful for slicing meat or fruit. After chewing is finished, the tongue manipulates the food into a bolus.
Once the bolus reaches the stomach and intestines, smooth muscle action continues to mechanically digest the bolus. Peristalsis and segmentation mix the bolus with gastric juices in the stomach and digestive juices in the small intestine. The result is a soupy mixture of food, water, and enzymes known as chyme. As this mixture moves to the large intestine, most of the water is recovered, leaving a drier waste material. Haustral contractions can move this waste slowly to help mix in bacteria. Reviewing Section 8.3, stimulation of involuntary muscle actions is controlled by the parasympathetic nervous system, the enteric nervous system, and some local hormones including gastrin. The sympathetic nervous system and local hormones, including secreting and CCK, controls inhibition of these muscles.
Do all animals use mechanical digestion? In some cases, no. For example, sponges lack a gastrointestinal system and are filter feeders: Sponges filter the water they live in, and feed on food particles that are much smaller than the sponge’s cells. With such tiny prey, sponges are unusual in that they rely on intracellular digestion.
Animals with a gastrovascular cavity (for example, flatworms), and tiny animals with a simple tubular gut may lack muscles in part or all of their digestive system. Some of these animals move their entire body to promote mechanical digestion, while others rely entirely on their mouthparts to perform mechanical digestion.
Unlike the obvious forces of mechanical digestion, chemical digestion happens at a microscopic scale, breaking the chemical bonds in food to release nutrients. Enzymes perform all chemical digestion in the gastrointestinal tract. You will recall that enzymes are proteins that act as catalysts to greatly accelerate specific chemical reactions. Most of the chemical reactions in chemical digestion involve the addition of a water molecule to a bond in an organic molecule. This is called hydrolysis (Greek, húdōr: water; lúsis: a loosening, setting free). Hydrolytic enzymes break large molecules (polymers) into smaller chemical units (monomers), allowing the monomers to be absorbed. For example, starches are polymers of glucose, proteins are polymers of amino acids, and DNA and RNA are polymers of nucleotides. The digestive system absorbs glucose, amino acids and nucleotides, not starches, proteins, and DNA. Even though lipids do not follow the classic polymer – monomer relationship, they still undergo a certain degree of hydrolysis.
Chemical digestion can occur within cells – intracellular digestion, or outside cells – extracellular digestion. In animals with a digestive system, most digestion is extracellular. This involves the secretion of hydrolytic enzymes into cavities of the digestive tract. Intracellular digestion is common in animals that lack digestive organs, for example the sponges. If you compare sponges with animals with a digestive system, why do you think sponges use intracellular digestion instead of extracellular digestion? Hint: think about sponges’ method of feeding[2].
When and where does chemical digestion occur? It can begin as soon as food in ingested. Remember your saliva contains hydrolytic enzymes, carbohydrases like amylase. Amylase will hydrolyze starches into glucose. Try holding a small piece of bread in your mouth for a few minutes. This should give amylase sufficient time to digest starch, release glucose, and the bread will begin to taste sweet. Oral amylase will be denatured by the stomach acids, and new carbohydrases will be released in the small intestine. There will be some lipases present within the saliva as well, to begin the digestion of lipids.
In most species, most of the chemical digestion is performed in the small intestine. The mucus of the oesophagus is for lubrication not digestion, and contrary to popular belief the stomach is not the main site of digestion, nor is digestion the role of stomach acids. There is a large group of mammals that break the “no digestion in the oesophagus rule”, the ruminants? Animals like cows, sheep, goats and deer have large folds and pouches in their oesophagus, creating three large chambers before the stomach. These chambers support billions of symbiotic microorganisms, including bacteria and protozoa, performing foregut fermentation. These microorganisms can digest tough plant fibres and release more nutrients for further processing.
If digestion mostly occurs in the small intestine, what happens in the stomach, what is the main function of the stomach? And why is it so acidic? The stomach itself is largely a food storage container. Since it can hold large quantities of food, there is a good chance many foreign microbes will be introduced into the body with the food. What do acids do to protein structure? Remember that many proteins will denature in the presence of acids. Stomach acids are the first line of defence against bacterial and viral proteins or enzymes. Additionally, the stomach releases proteases (enzymes that hydrolyse protein) to start protein digestion. Microbial proteins would be among the proteins digested.
The control over the stomach, its motility and secretions is complex. In review, the motility of the stomach is under the control of the CNS and PNS as well as a local enteric nervous system. There are also controls within the stomach itself (like gastrin) and controls from the small intestine (like secretin and CCK). It should not surprise you that the factors influencing stomach motility will also influence secretions and digestion within the stomach. Figure 8.9 shows a summary of the three main phases of stomach (gastric) secretion and the main controls over gastric secretions. Studying the names of the phases gives clues to the origin of influences over gastric secretion. The cephalic phase stimulates and inhibits gastric secretions via the CNS and PNS. The gastric phase includes controls originating from the stomach itself. Finally, the intestinal phase includes controls from the small intestine. Referring to both Figure 8 and Figure 8.10, the discussion will focus on the stomach’s production and inhibition two gastric secretions, HCl and pepsin (a protease).
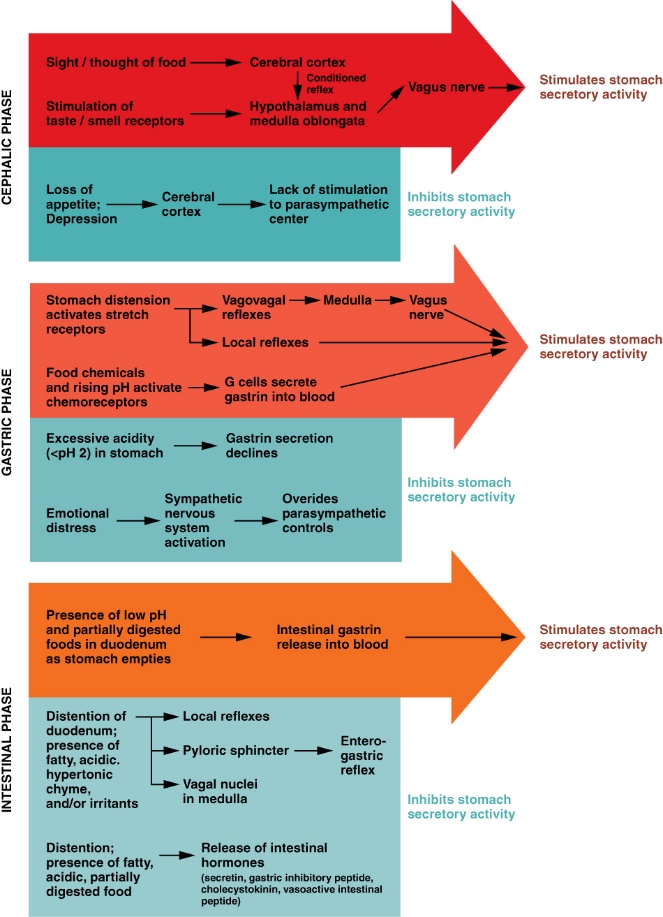
Figure 8.9 The three phases of gastric secretions, and their regulation. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
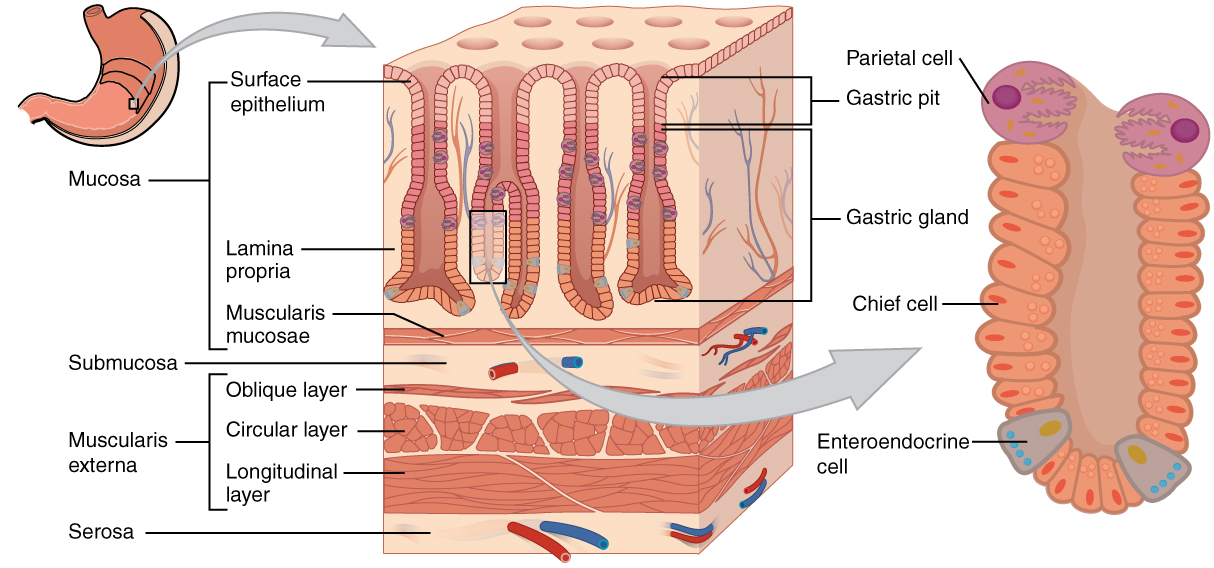
Figure 8.10 Histology of the human stomach, and the location of the primary secretory cells. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
Stimulation of gastric secretions begins during the cephalic phase. The sight, smell and taste of food results in a feed-forward response that stimulates HCl production from the parietal cells, pepsinogen production from the chief cells and gastrin from the enteroendocrine cells (Pepsinogen is an inactive precursor for the protein-hydrolyzing enzyme pepsin. What would happen if pepsin were produced in active form?) Many regions of the brain cause stimulation and this nervous stimulation will lead to the release of acetylcholine from the parasympathetic division, particularly from the vagus nerve. Once pepsinogen reaches the acidic conditions of the stomach cavity it is converted to pepsin. Many hydrolytic enzymes behave in this fashion, only becoming active in specific environments. This is serves a protective function for the secretory cells by limiting self-digestion.
During the gastric phase, decreasing pH (presence of more HCl) and increasing proteins in the stomach results in further release of gastrin from enteroendocrine cells. Gastrin stimulates the ENS, which overlaps the cephalic inputs, further increasing the output of HCl and pepsinogen. This is good example of a positive feedback loop, where one event reinforces the actions of another event, to move a system further and further from homeostasis. Gastrin will also stimulate the production of mucus to help protect the stomach lining from acids.
During the intestinal phase, presence of some acids and proteins in the duodenum (first portion of the small intestine) continues to stimulate the stomach through the release of intestinal gastrin. However, most gastric stimulation occurs prior to the intestinal phase.
Inhibition of gastric secretions will also involve all three phases. During the cephalic phase, many factors contribute to the lack of stimulation from the parasympathetic nervous system, so the focus of discussion will be events of the gastric and intestinal phases. During the gastric phase, a pH below 2 will inhibit gastrin release and limit the effects of the ENS. This also causes the release of somatostatin, which inhibits gastric secretions. Remember, stomach mucus production has been increasing as the pH drops. The mucus contains sodium bicarbonate; therefore, an increase in mucus concentration will buffer the pH, lowering the acid concentration. The loss of chyme and digested proteins to the duodenum reduces the protein stimulus; further reducing secretions. Finally, stress will activate the sympathetic division, which is antagonistic to the parasympathetic division. In contrast to the positive reinforcement between gastrin and pH, these mechanisms are components of a negative feedback system. Using one mechanism to counter the effect of another in a negative feedback loop is a common way to maintain homeostasis.
Despite all the inhibitory mechanisms in the stomach itself, the main inhibitory effects occur during the intestinal phase. Intestinal responses override many of the controls from the previous two phases. As gastric emptying proceeds, more chyme is moved from the stomach to the duodenum. Gastric emptying caused the duodenum stretch, the pH of the duodenum to drop, and an increase in concentration of proteins and fatty acids in the duodenum. An increase in gastric emptying leads to the inhibition of stomach processes, through the following actions by the duodenum:
- Stretching of the duodenum generates a reflex in the ENS which limits stomach motility and the secretion of gastric juices.
- Decrease in pH of the duodenum triggers a release of secretin from the duodenum, further reducing motility and secretion of gastric juices.
- Increase in fatty acids and proteins within the duodenum triggers a release of CCK from the duodenum, also reducing motility and gastric secretions.
There are numerous other signalling molecules released from both the stomach and/or the duodenum. Histamine, serotonin, ghrelin, motilin, GIP, and others all have impacts on digestion.
Now that the chyme is in the small intestine, pH must return to physiological neutral before enzymes can chemically digest the food. Accessory organs, most prominently the liver and pancreas, connect to the small intestine and provide secretions which buffer the pH and continue digestion (see Figure 8.11).
To review the events of gastric emptying: The release of secretin is stimulated by the introduction of acidic chyme to the small intestine. Once in the blood, secretin may target the stomach and reduce gastric secretions, thereby lowering the concentration of acids in the chyme. However, there are also secretin receptors in the liver and pancreas. Within these accessory organs, secretin stimulates the release of sodium bicarbonate, which completely neutralizes the acid chyme.
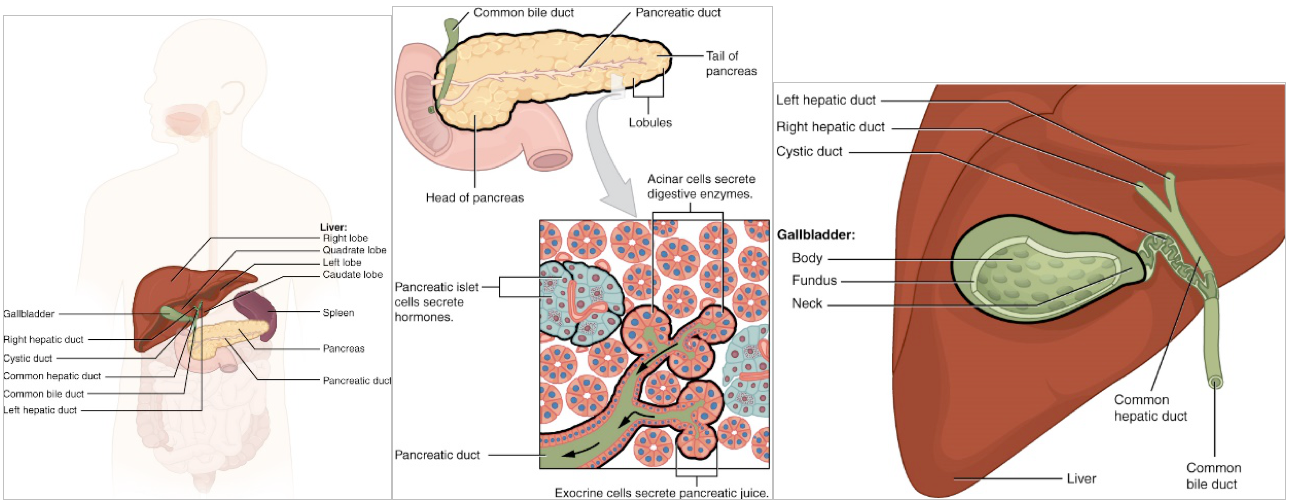 Figure 8.11 Human accessory organs involved in digestion. a) The location of the liver, pancreas and gall bladder, b) Position of the pancreas, location of the shared ducts with the liver, and histology demonstrating the endocrine and exocrine nature of the pancreas, c) The gall bladder and the shared ducts with the liver, including the common bile duct. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
Figure 8.11 Human accessory organs involved in digestion. a) The location of the liver, pancreas and gall bladder, b) Position of the pancreas, location of the shared ducts with the liver, and histology demonstrating the endocrine and exocrine nature of the pancreas, c) The gall bladder and the shared ducts with the liver, including the common bile duct. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
With the chyme returning to neutral pH, enzymes in the intestine can now begin extracellular digestion. The source of digestive enzymes is varied. Some enzymes will be released from intestinal glands, others from the epithelial lining of the intestine, yet others from the accessory organs. Recall the other local hormone, CCK. CCK is released when acid chyme, proteins and fatty acids enter the duodenum. CCK, like secretin, has target receptors in the accessory organs. CCK stimulates the release of bile from the liver, stored bile from the gall bladder and pancreatic enzymes from the exocrine portion of the pancreas. These digestive fluids will be released into the duodenum from the Sphincter of Oddi, a confluence of the common bile duct (draining the liver and gall bladder) and pancreatic ducts. We will now explore the role of bile and enzymes in the digestion of lipids, carbohydrates, proteins and nucleic acids.
Bile is produced by the liver and stored in the gall bladder. Unlike most of the other digestive fluids, bile doesn’t contain hydrolytic enzymes. Instead, it is a mixture of bile salts and other products like bilirubin (and other liver metabolites). Why are bile salts important? Remember that not all nutrients dissolve well in water. Many natural substances dissolve poorly in water because their electrostatic attraction to water is weak. These substances are called lipids. When you mix oil with water, the oil separates into large droplets. This is because water molecules don’t stick to oil molecules as strongly as water molecules stick to each other. So to digest lipids such as fats and oils, there needs to be a way to suspend these hydrophobic (Greek: húdōr, water; phóbos fear) materials in water with enough surface area. Bile acids are amphipathic, and therreby interact with both lipids and water. This helps suspend small lipid droplets in water for greater amounts of time, or helps create an emulsion. An emulsion is a mixture where one liquid is dispersed with, or suspended in, another liquid.
Following emulsification, a high surface area of contact between lipids and water allows access by enzymes called lipases, which carry out extracellular digestion of some of the lipids. Lipases are produced by the pancreas and small intestine. Triglycerides are the most abundant fats and oils digested by lipases. The lipase digests triglycerides by removing the outer two fatty acid chains from the glycerol backbone. The products of lipases are two fatty acid chains and a monoglyceride. Due to hydrophobic interactions, these products will form microscopic droplets known as micelles. Other fats, such as phospholipids and steroids, may not be chemically digested in the intestine lumen, but are absorbed unchanged.
Carbohydrates in the small intestine can be found as polysaccharides, disaccharides or monosaccharides, but only monosaccharides are absorbed (see Figure 11). A variety of carbohydrases will be responsible for generating the monosaccharides. Some common carbohydrases include amylase, lactase, maltase and sucrase. You have probably noticed the target of the enzyme is often incorporated into its name. Some of these enzymes are released by the pancreas (like amylase), but many are produced within the small intestine itself (like lactase, maltase and sucrase). The main sources of intestinal enzymes are epithelial cells, and the enzymes function along the absorptive surface known as the brush border. Explore Section 8.5, and Figure 8.15, for more information on the anatomy of the small intestine.
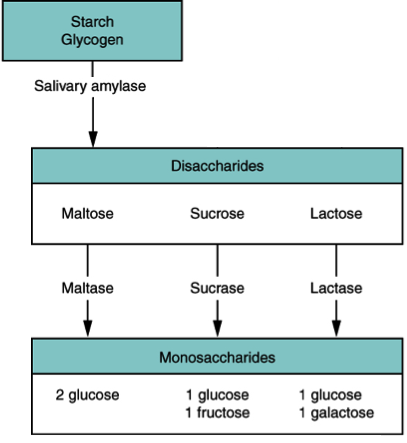
Figure 8.12 Carbohydrate digestion. Source: based on Donna Browne, Digestive System Module 7: Chemical Digestion and Absorption: A Closer Look, download for free at http://cnx.org/contents/3adcfc26-c4dc-463b-9c23-029d5ee258e8@1 CC BY 3.0 Rice University, 2014.
Protein digestion involves the digestion of any long polypeptide chains into smaller peptides, and eventually into individual amino acids, Figure 8.12. Recall protein digestion begins in the stomach with pepsin, but pepsin is only functional in low pH environments. New proteases and peptidases must be released within the small intestine that function at neutral pH. The pancreas produces many of these enzymes including trypsin, chymotrypsin, and carboxypeptidases. Others like enteropeptidase and aminopeptidases are produced with the small intestine. Just like pepsin, many of the proteases are released in an inactive state, known as a zymogen. Remember, this prevents these enzymes from degrading the proteins within the secreting cell. All three pancreatic proteases are released as zymogens: trypsinogen, chymotrypsinogen and procarboxypeptidase. One of the important roles of enteropeptidase is to activate trypsin within the small intestine. Once trypsin is active, it will activate more trypsin, as well as chymotrypsin, and carboxypeptidase.
How do these proteases digest proteins? Aminopeptidases and carboxypeptides are non-specific and digest any exposed amino acid from the end of a polypeptide chain. Remember a polypeptide is a directional molecule, with an amino-terminus and a carboxy-terminus. Even though these two enzymes are not specific for amino acids, can you guess where aminopeptidases and carboxypeptides are likely to function? Trypsin and chymotrypsin act differently, these enzymes will break large polypeptides into short peptides by looking for specific amino acids within the chain. Trypsin mostly identifies large positively charged amino acids like arginine and lysine, while chymotrypsin mostly targets large aromatic amino acids like phenylalanine and tryptophan.
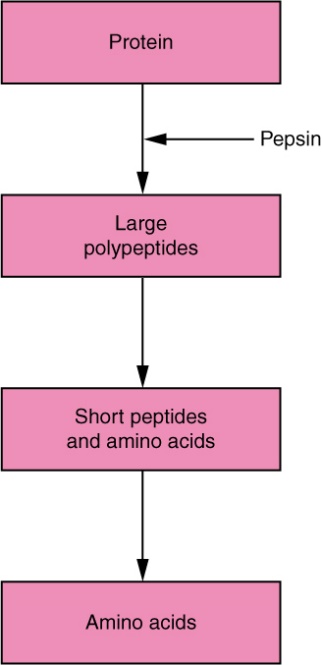
Figure 8.13 Protein digestion. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
Nucleic acids make up a very small portion of the diet, but are hugely important for cell function (DNA, RNA and ATP). DNA and RNA must be digested into the individual purines and pyrimidines. DNA is digested by deoxyribonucleases, and RNA is digested by ribonucleases. However, even individual purines and pyrimidines are too chemically complex to be absorbed, so brush border enzymes like nucleosidases and phosphatases degrade these into their constitutive parts (pentose sugars, nitrogen bases and phosphates).
The chyme and any remaining nutrients are slowly moved to the large intestine where the final stages of digestion occur. To reiterate, the small intestine is the main location for digestion, so the majority of enzyme secretion occurs in the small intestine. Most chemical activity of the large intestine is due to the gut microflora, composed mostly of bacteria. The bacteria are responsible for generating many vitamins and digestion of remaining carbohydrates, like fibre. Recent research has shown that gut microflora may even generate signalling molecules which alter the function of the digestive tract, and possibly even our eating behaviours.
What about water, minerals, electrolytes and vitamins? They don’t require chemical digestion, but they do need to be absorbed, as discussed below in Section 5.
As a summary of digestion, please review Figure 13.
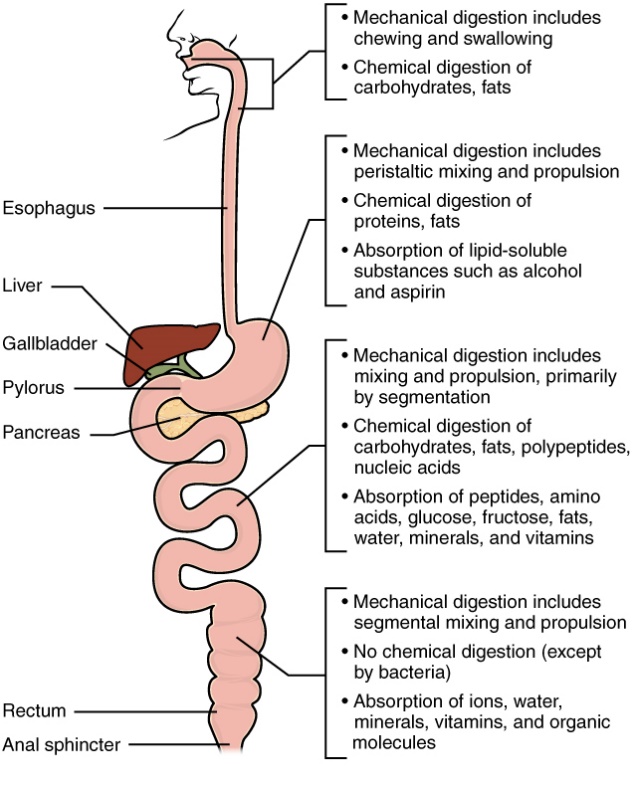
Figure 8.14 Human digestive system, with major locations of digestion and absorption. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
8.5 Absorption and Assimilation
After nutrients are digested, they should be absorbed and assimilated. Absorption is the uptake and transport of nutrients from the digested food to the blood or lymph. Columnar epithelial cells – densely pack rectangular shaped cells – line the stomach and intestines and are largely responsible for absorption. When studying the histology of the stomach and intestines, it is hard to ignore that this epithelial lining is just one cell deep. One might think that absorption is completely unregulated, as nutrients simply pour through this thin layer into the blood. However, research shows all nutrients, including water, must pass through the epithelial cells in a process known as transepithelial transport. Furthermore, tight junctions create an impermeable barrier between the epithelial cells, re-enforcing transepithelial transport. Why must absorption occur in this fashion? Quite simply, homeostasis. The body must be able to regulate nutrient uptake, forcing the passage of nutrients through cells and their membranes will contribute to this regulation.
Following absorption, assimilation can occur. Assimilation is the uptake of absorbed nutrients from the blood or lymph. This may be referred to as post-absorption by some researchers. Some assimilation processes will be mentioned here, but it does begin to overlap with energy balance in the body. As such, assimilation will be discussed in greater detail in following chapters.
The small intestine is the critical site for digestion, so it stands to reason it is also a critical site of absorption. Studying Figure 14, it should be clear why the small intestine is so important for absorption. There is a tremendous amount of surface area in the small intestine, due to several levels of folding. The wall of the small intestine forms large circular folds, almost doubling the inner surface area compared with a straight tube. The circular folds are covered by thin, fingerlike projections called villi (villus, singular). Finally, membranes of the absorptive epithelial cells are highly folded on a microscopic level, and project into the digestive cavity as microvilli; the collection of microvilli forms the brush border. What would you guess is the area of the absorptive surface of human small intestine? How many square metres? Many scientists estimate the surface area of the absorptive small intestine to be 250 m2, or approximately that of a tennis court!
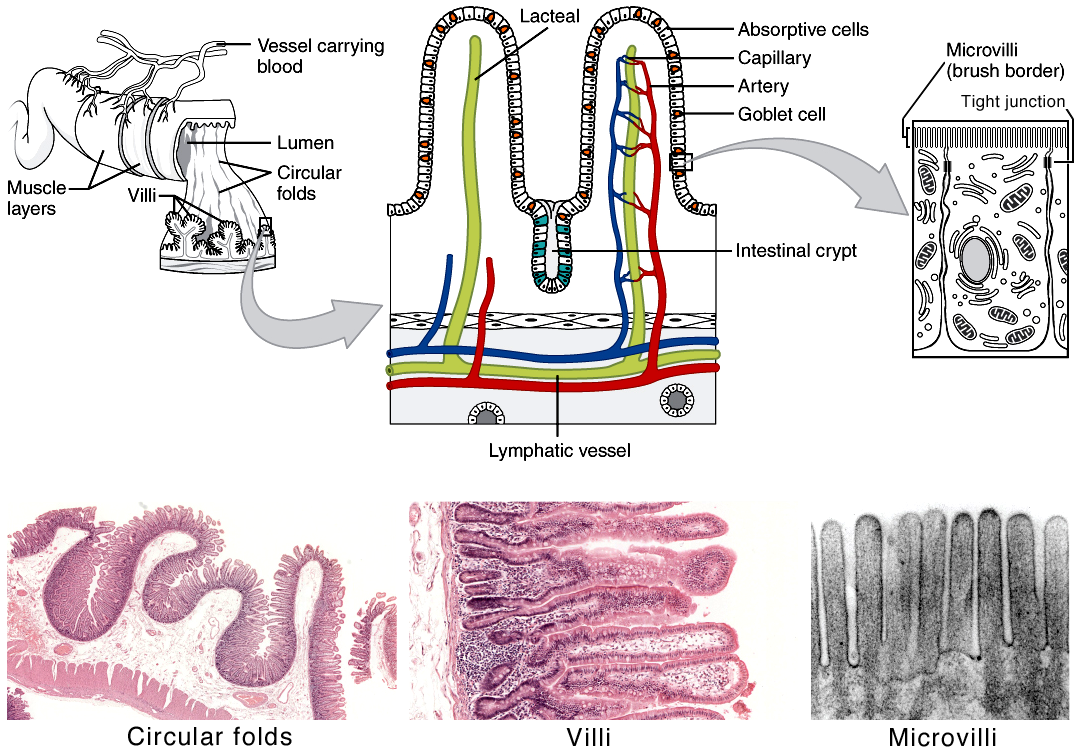
Figure 8.15 The small intestine has three levels of folding. Source: Adapted from Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
How are the different nutrients absorbed? Beginning with lipids, their absorption is largely passive transport or simple diffusion. Since membranes are lipids, digested lipids can move across epithelial membranes with ease. The creation of micelles is key to lipid absorption. The micelles are small enough to associate directly with the brush border. As they spin and bump into the brush border, individual fatty acids and monoglycerides will be released from the micelle and absorbed. Any phospholipids, steroids or fat soluble vitamins (vitamins A, K, D or E) associated with the micelle also diffuse into the epithelial cells. Within the cells, triglycerides are reassembled by the smooth ER, and then converted into protein coated fat droplets know as chylomicrons. Chylomicrons are then exported (bulk transported) to special lymph vessels within the villi, called lacteals. The creation and export of chylomicrons lowers the concentration of lipids within the cell, and ensures the transmembrane gradient will favour the diffusion of lipids into the cell from the digestive cavity. The lymph system eventually releases the lipids into the circulatory system near the heart.
Absorption of most monosaccharides and amino acids occurs via secondary active transport. Secondary active transport builds a strong gradient of monosaccharides and amino acids within the epithelial cells, by coupling their movement to the facilitated transport of sodium into the cells. As an example, let’s consider the absorption of glucose. As carbohydrates are digested, the glucose gradient strengthens in the digestive cavity. Unfortunately, cytoplasmic concentrations of glucose are still higher than the digestive cavity concentrations, so passive transport will not add more glucose to the cytoplasm of epithelial cells. In contrast, sodium concentrations are higher outside the epithelial cells and lower in the cytoplasm. This is due to cells actively pumping sodium from the cytoplasm into the extracellular environment, and the presence of sodium in food. Problem: how to import more glucose into the epithelial cells when the cells already contain glucose. Solution: place sodium-glucose symporters on the brush border. These symporters couple the passive transport of sodium into the cell, with the active transport of glucose into the cells. Therefore, the concentration of glucose increases over time, or the glucose gradient within the cell strengthens. In summary, the movement of sodium with its gradient is energy that drives the movement of glucose against its gradient.
How do the monosaccharides and amino acids complete transepithelial transport and make their way into the circulatory system? Recall, the concentrations of nutrients are typically higher in the epithelial cells, and lower in surrounding tissues. Passive transport, particularly facilitated transport, from the epithelial cells will occur. For example, glucose has specific glucose transport (GLUT) proteins to facilitate its movement from the epithelial cells into the surrounding tissues.
What about other organic nutrients? What about water? What about vitamins, minerals and electrolytes in the diet? Some monosaccharides like fructose can be moved through facilitated transport into epithelial cells. Water moves by osmosis, which is dependant on the osmolarity or tonicity of surrounding tissues. If the epithelial cells are hypotonic compared to the chyme, water stays in the chyme. If the cells are hypertonic, then water moves into the cells. Epithelial cells can change their tonicity to either increase or decrease water absorption. As previously mentioned, fat-soluble vitamins follow the lipid pathway. Some water-soluble vitamins can be facilitated, while others are actively transported. Some water-soluble vitamins can be absorbed through the entire intestinal system, while others are only absorbed in the small intestine. Minerals and electrolytes are also highly variable. Some are facilitated, while others like potassium (K+) and calcium (Ca2+) may require active transport and sodium antiporters. Since calcium is important for nerve function, muscle function and bone development, its absorption will be discussed in more detail.
The uptake of calcium (Ca2+) by the duodenum requires the presence of Vitamin D. Vitamin D is constructed from cholesterol, and processed by many regions in the body (including the liver and kidneys). One form of processed Vitamin D (calcitriol) is a steroid hormone that targets the absorptive cells of the duodenum. Steroid hormones can directly impact gene regulation, and in this case the epithelial cells immediately begin translation of calcium-binding membrane proteins. These calbindin proteins facilitate the uptake of calcium, and may require ATP to actively move calcium. Do you now understand why Vitamin D is added to milk? Beyond the duodenum, calcium can be absorbed passively into the epithelial cells depending on concentrations within the chyme. Absorption in the lower intestinal tract does not require the presence of Vitamin D. The movement of calcium into other tissues from the gut may require Vitamin D and calbindin proteins, or sodium antiporters. Calcium regulation within the kidney shares similar pathways to the digestive system. Uptake of calcium within gut is one small component of the calcium regulation pathway in the body, which involves the parathyroid, thyroid, bones, muscle and kidneys.
By the time the chyme reaches the last part of the small intestine, the small intestine wall has removed most of the available nutrients. The remaining material passes on to the large intestine through the ileocecal valve, a ring of muscle that closes firmly when the large intestine is stretched, preventing material from the large intestine from flowing back into the small intestine.
Can you guess what role the large intestine has in absorption? Hint: what substance is the single most important molecule for digestion, without which no digestion could take place?
Answer: One of the most important functions of the large intestine is to absorb water from the waste. Remember, food enters the small intestine as a wet, runny slurry called “chyme”. The small intestine eventually reabsorbs most of this water, but enough water remains that an animal would suffer dehydration if that water is lost from the body. The lining of the large intestine takes up sodium ions from the waste, and water follows by passive diffusion due to the osmotic gradient. Most of the remaining water is absorbed, resulting in semi-solid feces. Figure 15 shows a tremendous volume of water moving through the digestive tract, but very little is lost in the feces.
The large intestine is also responsible for recovering nutrients from the huge colony of bacteria fermenting the food remnants in the large intestine before defecation. Many vitamins are recovered here, both from the waste and from the metabolism of the microflora. Any remaining electrolytes and minerals will be regulated, largely based on concentration gradients. For instance, if there is still a high concentration of calcium in the waste and if the body requires more, calcium will be absorbed via passive transport.
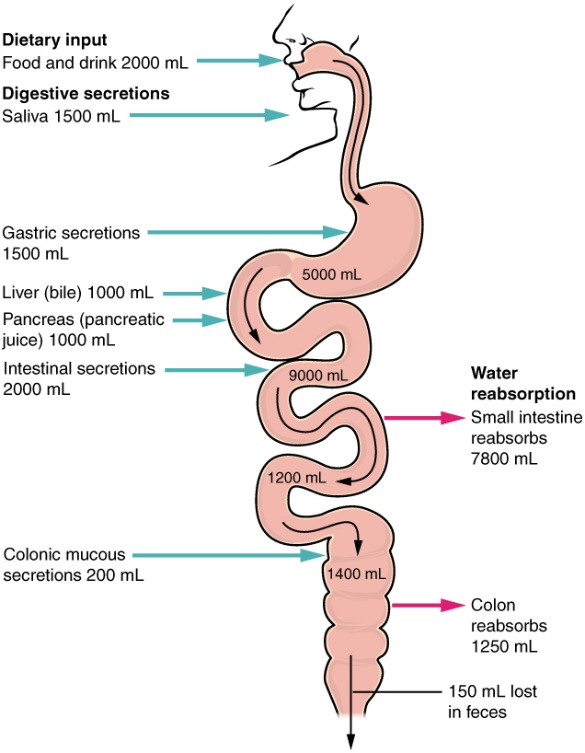
Figure 8.16 Ingestion, usage and loss of water within the human gastrointestinal system. Source: Anatomy & Physiology, download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@6.27. CC BY 3.0 Rice University, 2014.
Some organisms rely on the large intestine microflora more than others. Horses and rabbits are hind-gut fermenters. Contrary to fore-gut fermenters, horse and rabbits rely on bacteria in their large intestines to break down fibre and release even more nutrients from the waste. Some species of rabbit and the occasional gorilla, may even eat the feces to recover the nutrients their large intestine was unable to absorb. What an odd method of nutrient translocation!
Assimilation is the uptake of nutrients from the blood and lymph. All the blood from the digestive system flows through the liver before being moved to through the heart and to the rest of the body. That means monosaccharides and amino acids all move through the liver before reaching the rest of the body. Remember lipids use the lymphatic system and reach the heart before the liver. The focus of discussion will be three major organic nutrients: glucose, amino acids and triglycerides. Below is a summary of major sites of assimilation and general functions:
- Glucose – Initially assimilated by the liver, but muscle and nervous tissues use large quantities. It is a primary energy source, can be structural.
- Amino acids – Assimilated by all tissues, liver and muscle are common. Construction of proteins and enzymes, structural and metabolism.
- Triglycerides – Assimilated by adipose tissue and muscle, later the liver. Secondary energy source, structural and thermoregulation.
Glucose is one of the primary energy sources in the body. Nervous tissues have a particularly high absolute need for glucose, and have highest priority when glucose is limited. The liver and muscle tissues have a lower affinity for glucose, but are major sites of glucose uptake and storage when the nervous system has enough. However, assimilation of glucose by the liver and muscle can change depending on the quantities glucose in the blood and the presence or absence of the pancreatic hormone, insulin. The regulation of glucose assimilation by liver and muscle tissues is as follows:
- Post-absorption, glucose levels in the blood will be high.
- The pancreas will respond to that stimulus and to input from the hypothalamus.
- Beta-cells within the pancreas produce and release insulin.
- Insulin increases the sensitivity of muscle and liver tissues to glucose.
- More glucose is assimilated by the liver and muscle. In the liver, it is transformed into glycogen.
- Blood glucose levels drop, insulin production goes down, assimilation slows.
Those events are one half of a negative feedback pathway controlling blood glucose levels. The second half of the pathway does not involve digestion and assimilation in terms of the digestive system, but are important processes nonetheless:
- Blood glucose levels drop.
- Alpha-cells of the pancreas produce glucagon.
- Glucagon targets the liver and initiates glycogen digestion (Glycogenolysis).
- More glucose is released into the blood.
- Alpha-cells and glucagon are inhibited, Beta-cells are stimulated.
Amino acids are important for the creation of proteins, and are assimilated by any living cell. Amino acids can be used as an energy source, but this typically occurs after carbohydrates and lipid stores have been depleted. After triglycerides travel through the lymphatic system, they are released into circulatory system near the heart. Because triglycerides are hydrophobic, they do not dissolve in the blood. Instead, circulating triglycerides are chaperoned by proteins. Triglyceride assimilation will once again require lipases to digest them into fatty acids and monoglycerides. This digestion in the blood often occurs prior to assimilation by adipose (fat) tissue, muscles, or the liver.
8.6 Elimination of Wastes
The large intestine (the colon and rectum) is responsible for compaction and removal of waste materials, now called feces. Recall haustral contractions slowly move the feces through the large intestine toward the anus. Along the way, some digestion and recovery of remaining nutrients occurs. When new food is ingested and begins to move through the digestive system, mass movements may replace haustral contractions. These can move feces very quickly through the colon toward the rectum for storage. If large amounts of feces build up in the rectum, stretching of the rectum generates a reflex, known as the defecation reflex. During this process, the internal anal sphincter relaxes and the large intestine forcefully contracts, moving feces into the anal canal. Defecation may not immediately occur because the external anal sphincter (commonly called the anus) is under voluntary control. Eventually, enough pressure will build up in the anal canal and the feces will be released, or egested. Quite literally, we have reached the end of this chapter!
8.7 Summary
- Food is a source of energy, and a source of chemical building blocks for the animal body.
- The body can only make use of food if the food is broken down into simpler chemicals.
- This is because the body can only absorb and use simple, small molecules.
- Chemical digestion uses hydrolysis to break down the large, complex molecules of food into smaller, simpler molecules.
- Proteins called digestive enzymes act as catalysts to hydrolyse food molecules.
- Chemical digestion can only happen if a large surface area of food is exposed to water and digestive enzymes.
- Mechanical digestion by the teeth, tongue, and stomach grinds solid food to increase the surface area of food exposed to digestive juices.
- Acid in the stomach helps soften food for mechanical digestion, and kills most microbes.
- The stomach also adds a large amount of water to the food, and the food enters the small intestine from the stomach as a runny, watery slurry called “chyme.”
- As soon as the chyme flows into the small intestine, it mixes with juice from the common bile duct
- Muscle-powered movement in the digestive tract includes peristalsis, segmentation, and haustral contractions.
- Muscular valves called sphincters control the flow of material from one part of the digestive tract to another.
8.8 Suggested Readings
8.9 Glossary
Alimentary canal – The tube or pipe that goes from your mouth to your anus.
Amylase – Enzyme that hydrolyses starch and glycogen.
Bile – Liquid that the liver secretes into the bile duct. Contains bile salts.
Bile acid or bile salt – Amphipathic compounds secreted in bile. Responsible for emulsification of lipids.
Bolus – Lump of material passed in one direction, usually by peristalsis.
Brush border – The dense lawn of microvilli responsible for the absorption of nutrients. In cross-section, looks like the bristles of a brush. See microvillus.
Carboxypeptidase – Protease secreted by pancreas, works at the exposed carboxyl end of a protein.
CCK – Cholecystokinin
Cholecystokinin – Hormone released by the small intestine. Regulates stomach action, stimulates digestive secretions from liver and pancreas, reduces appetite.
Chyle – Chylomicron-rich lymph, taken up and carried by lacteals in the villi of the small intestine. Not to be confused with chyme.
Chylomicron – Droplet of newly digested and absorbed lipids in chyle and blood. Stabilised by specific proteins on its surface. Made by the absorptive cells of the small intestine.
Chyme – Soupy mixture of food with digestive juices.
CNS – Central nervous system.
Dietary fibre – Indigestible components of food.
Colon – Main portion of large intestine, site of bacterial fermentation, and absorbs water, minerals, and some vitamins from waste material.
Duodenum – First portion of the small intestine. It is where the common bile duct and the pancreatic ducts empty into the intestine. Active in chemical digestion.
Emulsion – Mixture of two or more liquids.
Egestion – Expulsion of food waste.
ENS – Enteric nervous system.
Esophagus – See oesophagus.
Facial nerve – Seventh cranial nerve, connects to anterior two thirds of tongue and carries taste signals.
Feces – Dry waste material containing dietary fibre and other undigested material, bacteria, and waste products.
Fibre – See dietary fibre.
Gall bladder – Bile storage organ surrounded by the liver.
Gastrin – Hormone produced by the stomach and small intestine. Stimulates HCl production by stomach, and motility of stomach and intestines.
Gastrointestinal tract – Tubular digestive system. See alimentary canal.
Gastronomy – Like astronomy, only you’re exploring the universe of well-prepared foods.
Gastrovascular cavity – Digestive cavity within some animals with only one entrance, the mouth. Can play circulatory and supportive roles in addition to digestion.
Glossopharyngeal nerve – Ninth cranial nerve, from posterior third of tongue, important in swallowing reflex.
Haustra – Large pockets in the large intestine, containing chyme or waste.
Haustral contractions – Slow rhythmic movements, mixing chyme or waste in the large intestine.
Hepatic portal vein – Blood vessel that carries blood loaded with water-soluble nutrients directly from the small intestine to the liver, resulting in first-pass metabolism of substances absorbed from the food.
Heterotrophic – Getting energy and carbon from organic molecules from other living things.
Hydrolysis – Cutting a molecule into two smaller molecules by adding a molecule of water. Hydrolysis is the chemical reaction that digests carbohydrates, proteins, and fats into the smaller, simpler chemicals the body can absorb.
Hydrophobic – Repels water because binding to water molecules is weak.
Lacteal – Vessel of the lymphatic system, leading from a villus in the intestine wall.
Larynx – Region in trachea containing vocal cords.
Lipase – Lipid digesting enzyme.
Lipid – Any natural substance that dissolves poorly in water because it binds only weakly to water molecules. Part or all of a lipid molecule is hydrophobic. Fats, oils, and sterols are examples of lipids.
Liver – Accessory digestive organ. Produces bile and bile salts, detoxifies many compounds, stores glycogen, and performs many additional metabolic functions unrelated to digestion.
Lumen – The space inside a tube-like or bag-like organ.
Micelle – Tiny droplet of lipid, only a few molecules wide, small enough that lipid molecules exchange rapidly between the droplet and the surrounding water. Formation requires a surfactant.
Microflora – The colony of bacteria and other microbes associated with your body. Mainly symbiotic bacteria in the digestive tract, helping digestion.
Microvillus – Thin extension of the plasma membrane, stiffened on he inside by a bundle of actin microfilaments. The lumen-facing side of epithelial cells lining the small intestine is packed with microvilli.
Mucus – Watery, slimy secretion that lubricates the inner surfaces of the digestive tract.
Nutrient – Any substance, other than gases, needed by an organism to maintain metabolism, growth, and homeostasis.
Oesophagus – Muscular tube running from the pharynx to the stomach.
Pancreas – An exocrine gland, secreting bicarbonate and digestive enzymes into the pancreatic ducts, and an endocrine gland, secreting the hormones insulin anf glucagon.
Pepsin – Protease found in the stomach, active below pH 4.
Peristalsis – Unidirectional waves of contractions moving food through the digestive system.
Pharynx – Troat region where the mouth, nasal cavity, larynx, and oesophagus converge.
PNS – Peripheral nervous system.
Protease – Any enzyme that hydrolyzes polypeptides (proteins).
Proenzyme – An enzyme before it has been activated. Alternatively called a zymogen.
Rumen – Large bulge in the oesophagus in some herbivores, housing symbiotic bacteria able to digest cellulose from plant cell walls.
Saliva – Watery secretion in mouth. Lubricates food, helps mechanical digestion, and contains some digestive enzymes (in humans, amylase and lipase).
Salivary glands – Exocrine glands around the oral cavity producing saliva.
Secretin – Hormone released by the small intestine, slows stomach activity and stimulates the pancreas and liver to release bicarbonate.
Segmentation – Mixing movements in the small intestine.
Surfactant – Substances that lower the surface tension of liquids, increasing mixing and emulsification.
Sphincter – A muscular valve that controls the flow of material from one part of a tubular organ system to another.
Stomach – Organ that stores swallowed food. Produces acid and performs some protein digestion.
Trypsin – Abundant protease released by the pancreas into the small intestine.
Villus – Finger-like folds of the inner wall of the small intestine.
Zymogen – See proenzyme.
- Inorganic = water to zinc in the list, many of these are the minerals in your diet, ions are often called electrolytes; Organic = glucose to niacin in the list; Four main categories of organics: carbohydrates, proteins, nucleic acids and lipids ↵
- Since sponges are filter feeders, and pump massive amounts of water through their bodies, extracellular digestion could result in nutrient loss to the environment. ↵
