1. The Animal Kingdom
Chapter 1 – Evolutionary Aspects of the Animal Kingdom
- 1.1 Introduction
- 1.2 Common Features of Animals
- 1.3 Embryonic Development
- 1.4 The Origin of Animals
- 1.5 Gas Exchange and the Evolution of Animals
- 1.6 Cell Interactions and Tissues
- 1.7 Animal Tissues
- 1.8 Animal Body Plan
- 1.9 The Chordates
- 1.10 Summary
- 1.11 Suggested Readings
- 1.12 Glossary
1.1 Introduction
The word “animal” came from the Latin word animalis, meaning with “breath” (anima) or “soul”, as seen in the Latin Bible[1]. People often use the word “animal” to refer to non-human species only, but the biological definition refers to all members of the kingdom Animalia, which includes creatures as diverse as sponges, jellyfish, insects, and humans. The kingdom Animalia is also known as Metazoa (Greek, meta: along with; zôion: animal).
In the past, researchers divided Metazoa into two broad groups:
- Some animals have a simple level of organisation, with relatively few cell types and no distinct organs. These simple animals were grouped in the subkingdom Parazoa (Greek, para: at one side of). The sponges are the most common example of animals with such a simple body plan.
- Animals with more complex bodies were traditionally termed Eumetazoa (Greek: eús: good or true). A distinguishing feature of Eumetazoa is the high level of cell differentiation, with cells organized into specialised tissues, such as nervous, muscular, epithelial, and connective tissues.
As researchers have gained a better understanding of the diversity of animals in recent years, they realised that one or more unusual groups of animals may have lost complex tissues and organs during evolution. The blob-like Placozoa might be an example of an animal group that’s greatly simplified compared with its ancestors. Researchers also realised that one or more groups of animals may have independently evolved complex tissues and organs. The comb jellies (Ctenophores) might be an example of an animal group that evolved muscles, a nervous system, sensory organs, and a digestive tract independently of other animals. Since “Parazoa” and “Eumetazoa” no longer seem to be natural groups of animals, these two terms aren’t used much anymore.
1.2 Common Features of Animals
There are more than one million known animal species on earth, ranging from ants to zebras. Given that animals have evolved to adapt to environmental conditions as diverse as deep sea, rain forest, and deserts, it should be no surprise that animals show a wide array of different morphological (body shape and structure) and physiological (body function) characteristics. However, most animals have several characteristics in common:
Animals Reproduce – Like any living thing, all animals need to reproduce in order for the species to survive. Most animal species have male and female individuals that reproduce sexually by fusing the small gamete, sperm, with the large gamete, egg. Gametes carrying half of the chromosomal complement of the somatic cells are produced by meiotic cell division in the animal’s gonads. The result of the fusion between the sperm and the egg is a new entity termed the zygote, which carries the full chromosome complement of the species. The zygote is the cell that develops into the new individual.
Thus, sexual reproduction produces individuals with genetic characteristics from both parents, ensuring a mixing of gene variants within a species.
Sexual reproduction does not always involve two separate sexes: Some species have individuals that possess both male and female reproductive organs. They are referred to as hermaphrodites[2].
Although most animals have the ability to reproduce sexually, many animal species reproduce asexually. Asexual reproduction doesn’t require the involvement of a second individual, and does not require fusion of gametes. Asexual reproduction produces genetically similar or identical copies of the parent (called clones). Some species can reproduce asexually by budding, or by splitting (fission), or by tissue fragmentation, and yet others can reproduce by parthenogenesis (Greek, parthenos: virgin, and genesis: creation), where an unfertilized egg develops into an embryo. The chapter on reproduction describes the various modes of reproduction in more detail.
Animals are Multicellular – Beyond the zygote stage of their life cycle, all animals are multicellular, meaning that the body is composed of many cells of different kinds, each cell type being specialised for a different role in the body. Multiple specialised cell types working together in a single body is a characteristic of all animals. As discussed above, all cells of the body are descendants of the zygote by cell division. As cell division the embryo creates more and more cells, the cells communicate with each other to decide which cells develop into which organs and cells types.
The development of differing cell types is called differentiation, and the formation of organs is called organogenesis.
Animals Eat – Animals are heterotrophs[3], meaning that they can’t fix carbon, and therefore can’t make organic molecules from inorganic sources to feed themselves. This contrasts with most plants, which capture energy from sunlight to fix carbon. Most animals must feed on other organisms. Eating provides energy and essential nutrients through ingestion of raw materials that are digested and reduced to metabolically usable molecules. These include simple sugars, amino acids and fatty acids that are metabolized to obtain energy. Based on their feeding habits, animals are subdivided into the following groups:
- Carnivores – diet mainly or exclusively animal food;
- Herbivores – diet mainly or exclusively vegetable food;
- Omnivores – diet both animal and vegetable food;
- Parasites – are animals that have a non-mutual symbiotic[4]
- Detritivores (also called saprophages) – diet on detritus composed of dead organic matter from animals or plants.
Animals Move – Movement differentiates animals from most plants and fungi. Most animals move – at least at some point in their lives. Movement requires complex behavioural responses to stimuli: With the exception of parazoans, animals have a sensitive system of nervous and muscle cells. Nerves and muscles allow animals to move in response to environmental signals, and facilitate spontaneous and coordinated movements to capture prey for food.
Animals Cells Do Not Have Cell Walls – The cells of plants, fungi, many signle-celled eukaryotes, and most prokaryotes have a solid outer covering, called the cell wall, outside the plasma membrane. Animal cells are different in that they lack a cell wall. The absence of a cell wall is a characteristic of all animals.
If animal cells don’t have a cell wall, how do animal cells get their shapes? The scaffold of internal proteins that compose the cytoskeleton supports cells internally.
Vertebrate animal bodies are supported by an internal skeleton (“endoskeleton”), composed of cartilage, such as the cartilaginous fishes, or mineralized connective tissue of the bones of most of the vertebrates. Many invertebrates have an external skeleton, or exoskeleton, that enclose and protect the soft tissues and organs of the body.
1.3 Embryonic Development
Zygotes undergo rapid mitotic cell divisions with no significant growth in mass (Figure 1.1). Development of the early embryo in most animals shows a pattern of three main stages:
- the morula,
- the blastula, and
- the gastrula.
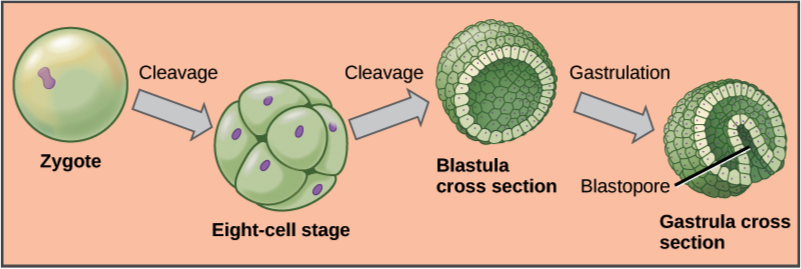
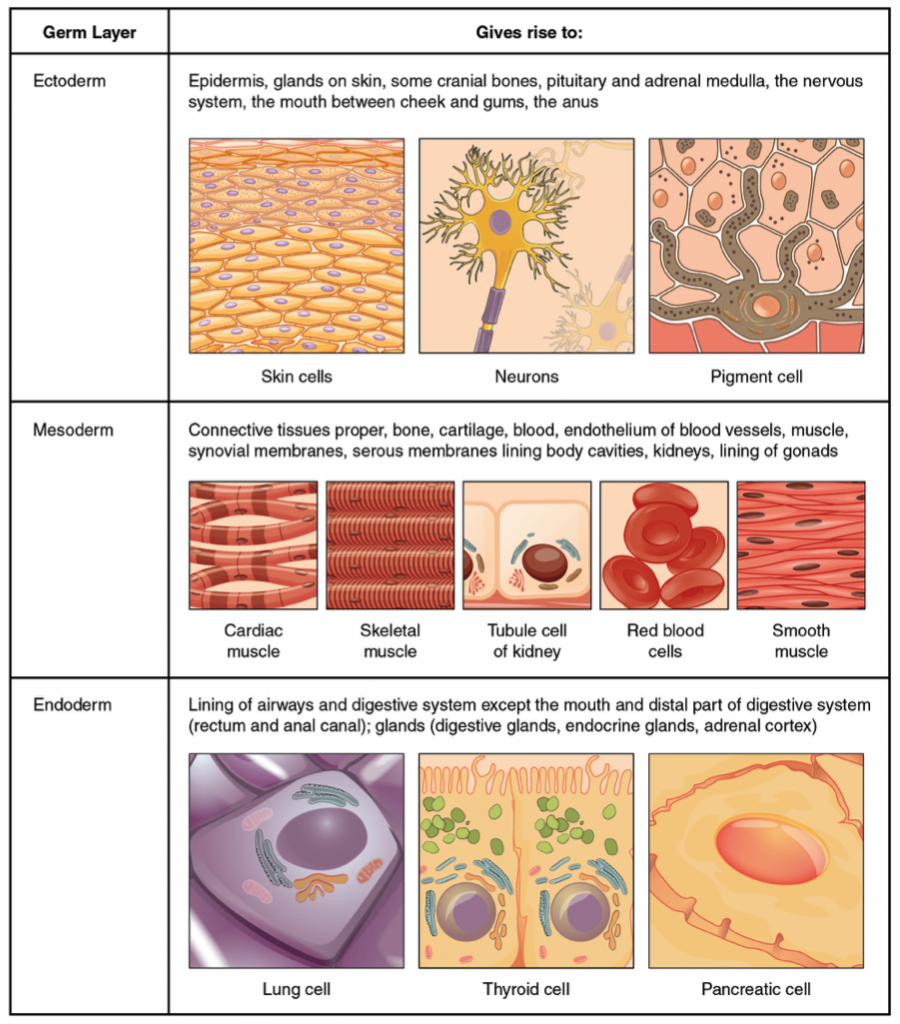
At the stage of 8 to 16 cells, the embryo forms a compact mass of cells termed a morula, because it resembles a mulberry (Latin, morus: mulberry; Latin, -ula: little). The morula develops into a hollow sphere of a single layer of cells, termed a blastula (Greek, blastós: sprout). As a blastula invaginates (a pocket folds inwards) and undergoes further cell growth, it differentiates into gastrula (Greek, gastḗr: stomach), a stage with three distinct layers of cells that give origin to the three primary tissues: ectoderm, mesoderm, and endoderm (Greek: ektos: outside; mesos: middle; endon: within; and derm: skin).
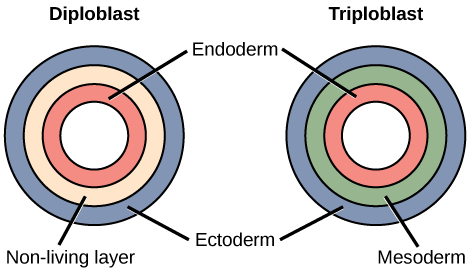
In the adult, these primitive germ layers differentiate to form tissues and organs. Some eumetazoans are termed diploblastic because the blastula generates only two primary germ layers, the ectoderm and the endoderm. Jellyfish, corals, hydra, and the unrelated comb jellies are examples of diploblastic animals.
Animals with tissues originating from three embryonic cell layers are termed triploblastic species. Table 1.1 summarizes the origins of adult tissue from the three primary germ layers.
| Primary Tissue Layer | Adult Tissues and Organs |
|---|---|
| Ectoderm | Skin and its derivation: hair, feathers, scales, and nails; nervous system and adrenal medulla; lens, retina and cornea of the eye; sweat and mammary glands. |
| Mesoderm | Muscles, bones and cartilage; circulatory system, internal reproductive organs, kidneys and outer wall of the digestive tract. |
| Endoderm | Lining of the digestive tract, liver, pancreas, lining of the respiratory tract, thyroid gland, lining of the urethra and urinary bladder. |
Let’s take a moment to look at embryo development in more detail.
The egg cell is covered with acellular and cellular layers of protective envelopes. The first layer of the egg envelope is the vitelline membrane, or zona pellucida in mammals. This layer is composed of glycoproteins that protect the egg and prevents attachment during it passage through the oviduct. The egg also contains all the cell components required for cell division and yolk, which provide the nutrients that support the initial stages of embryo development. Yolk is abundant in the eggs of insects, reptiles, and birds, which are laid in condition where there is no supply of nutrients. Little yolk is found in the eggs of species that provide nutrients through the placenta. Yolk can be concentrated at one end, in the centre or evenly distributed throughout, giving the egg of some species an asymmetric shape. In the eventual development, the animal pole gives rise to surface structures and to the anterior end of the embryo; while the vegetal pole gives rise to internal structures such as the gut, along the posterior end of the embryo. Egg polarity also plays a role in setting the body axes of bilaterally symmetrical animals.
Following the fusion of the egg with the sperm, the zygote enters in a cycle of rapid divisions, by cleavage into half-size daughter cells with no significant growth. These early embryonic cells, termed blastomeres, alternate between mitotic (M) phase and DNA synthesis (S) phase, with no growth phase (G1 or G2), resulting in smaller cells after each division. By the time the zygote divides into 16 to 32 blastomeres, it starts forming cell adhesions that hold the blastomeres together. These cell adhesions include desmosomes, tight junctions, and gap junctions – the same kinds of cell adhesions that are typical of epithelial cells. You’ll read more about these cell adhesions later in this chapter, in Section 1.6. During the zygote stage, blastomeres are autonomous cells, meaning that are not specified to be any particular tissue in the differentiation process.
Morula. The early zygote is now a compact mass of cells, called a morula because it resembles a mulberry (Latin, morus: mulberry; -ula: little). Two main events occur on the cell membrane at this stage of embryo development. In the first event, termed compaction, the 8-cell embryo expresses cell adhesion molecules (E-cadherin in mammals) that join cells together, creating a barrier that makes the embryo surface impermeable. and control paracellular transport. Thus, by compaction, the embryo becomes a solid ball of cells. The second main event is an increase in the expression of the Na+/K+-ATPase. This enzyme pumps Na+/ on one side of the cell, increasing osmotic pressure on the interior of the morula, leading to accumulation of fluid by osmosis. This forms a cavity inside the embryo.
Blastula. Impermeabilization of the compacted blastomeres during the morula stage and cavitation of the interior form a fluid-filled cavity called the blastocoel (also spelled blastocoele or blastocele). Thus, the morula become transformed into a hollow ball of cells that in mammals is referred to as a blastocyst (Greek, blastós: a sprout, kystis: bladder). The blastocyst holds a group of primordial pluripotent cells, called the inner cell mass (ICM), that subsequently forms the embryo. During the blastula stage the early embryo start to establish cell polarity, cell specification, axis formation, and regulation of gene expression. The tissue extending perpendicularly to the animal-vegetal axis creates a small opening in the blastula, termed blastopore, that invaginate forming the archenteron, which is a cavity within the blastocoel lined with endoderm and sealed with a yolk-rich tissue that provide nutrients. The archenteron will become the animal’s digestive tract, which extends, shaping a cylindrical structure that touches the ectoderm, forming the mouth; While the blastopore, sealed with yolk tissue, becomes the anus. In humans, blastocyst formation begins about 5 days after fertilization when morula reach the stage of 16 cells.
Gastrula. From the first zygote division to the stage of blastula the embryo is just a continuous epithelial sheet of cells. Blastula folds in on itself and, about seven days after fertilization in humans, the embryo enters in rapid rounds of mitosis. A strip on the surface of blastula, termed the primitive streak, determines the site of gastrulation. At this point epithelial cells undergo a transition to mesenchymal cells that ingress into the blastula through the primitive streak. Then cell differentiation starts and the embryo establishes distinct cell lineages, set up the basic axes of the body – dorsal-ventral, anterior-posterior – and internalize different cell types including those of the primitive gut system, and the primordial germ cells that migrate via the gut to the developing gonads where undergo meiotic division. By the end of gastrulation, the embryo of triploblastic organism is organized into well-defined multilayered structures composed of the outer layer, ectoderm (ecto = outside; derm = skin), the middle layer mesoderm (meso = middle), and the inner layers endoderm (endo = inside). The gastrula of diploblastic organisms – Cnidaria and Ctenophora – has only ectoderm and endoderm, also termed hypoblast and epiblast, respectively. By the time the embryo reaches about 200–300 cells and a diameter of about 0.1–0.2 mm, it undergoes implantation into the endometrium of the uterine wall. Embedding of the blastocyst into the endometrium requires that it hatch from the zona pellucida to start implantation into the uterus. The trophectoderm differentiates into the trophoblast, which secretes proteolytic enzymes that break down the extracellular matrix of the lining of the uterus (endometrium) and extends finger-like projections invading the maternal tissue. The trophoblast is not part of the embryo itself, but form part of the extraembryonic membranes that supports embryo growth, provide nourishment and develops into a large part of the placenta.
Differentiation and organogenesis. The process of differentiation is regulated by specific chemical signals from the cell’s embryonic environment that sets in play a cascade of events that regulates gene expression. Embryonic stem cells express specific genes that will determine their ultimate cell type. For example, some cells in the ectoderm will express the genes specific to skin cells. As a result, these cells will take on the shape and characteristics of epidermal cells. Some differentiated cells continue dividing through the life of the individuals, while in others differentiation is terminal, such as in the erythropoietic tissue, striated muscle epidermis and intestinal cells.
The first evidence of differentiation is observed during gastrulation, when the rapidly growing mesoderm gives rise to a flexible rod of cartilage-like tissue, termed the notochord. The notochord produces signalling proteins, including bone morphogenetic proteins and cadherins that target the ectoderm. In response the ectoderm differentiates into the neural plate, a primitive neural structure composed of neuroepithelial cells that diversify into immature neurons and neuroglia cells. The neural plate will become the dorsal midline of the body along and above the notochord. At both sides the neural plate a neural crest border in continuity with the rest of the ectoderm. The entire process, is termed primary neurulation that can be described in a sequence of events as follows:
- The neural plate folds, forming the neural groove. When the two ends of the neural groove join at the neural crest separates from the ectoderm forming a tubular structure termed neural tube;
- The neural tube gives origin to the neurons of the central nervous system, while the cells of the neural crest differentiate forming most of the tissues of the peripheral nervous system;
- On both sides along the notochord, cells of the neural crest form the ganglia of the spinal chain. The neural crest cells also differentiate into melanocytes, the melanin-producing cells of the bottom layer of the epidermis, the middle layer of the eye, the inner ear, vaginal epithelium, meninges, bones, and heart. Melanocytes produce melanin, a dark pigment primarily responsible for skin colour.
- The presence of the notochord defines the phylum Chordata. In most chordates, the notochord degenerates by the time the animal is an adult. In vertebrate animals, the notochord serves as a precursor for the spinal column, and forms the sites for muscle attachment. In the spinal column, the notochord persists as jelly-like material of collagen fibres, termed the nucleus pulposus, that forms the inner core of the vertebral disc. This elastic inner structure allows the vertebral disc to withstand forces of compression and torsion. In other words, it’s a shock absorber.
- The rest of the mesoderm derives into paraxial mesoderm, also termed somitic mesoderm, a tissue that flanks and forms simultaneously with the neural tube. The paraxial mesoderm give rise to a block of tissue, termed somites, that form muscle and tissues of the back part of the body, including connective tissue and the dermis.
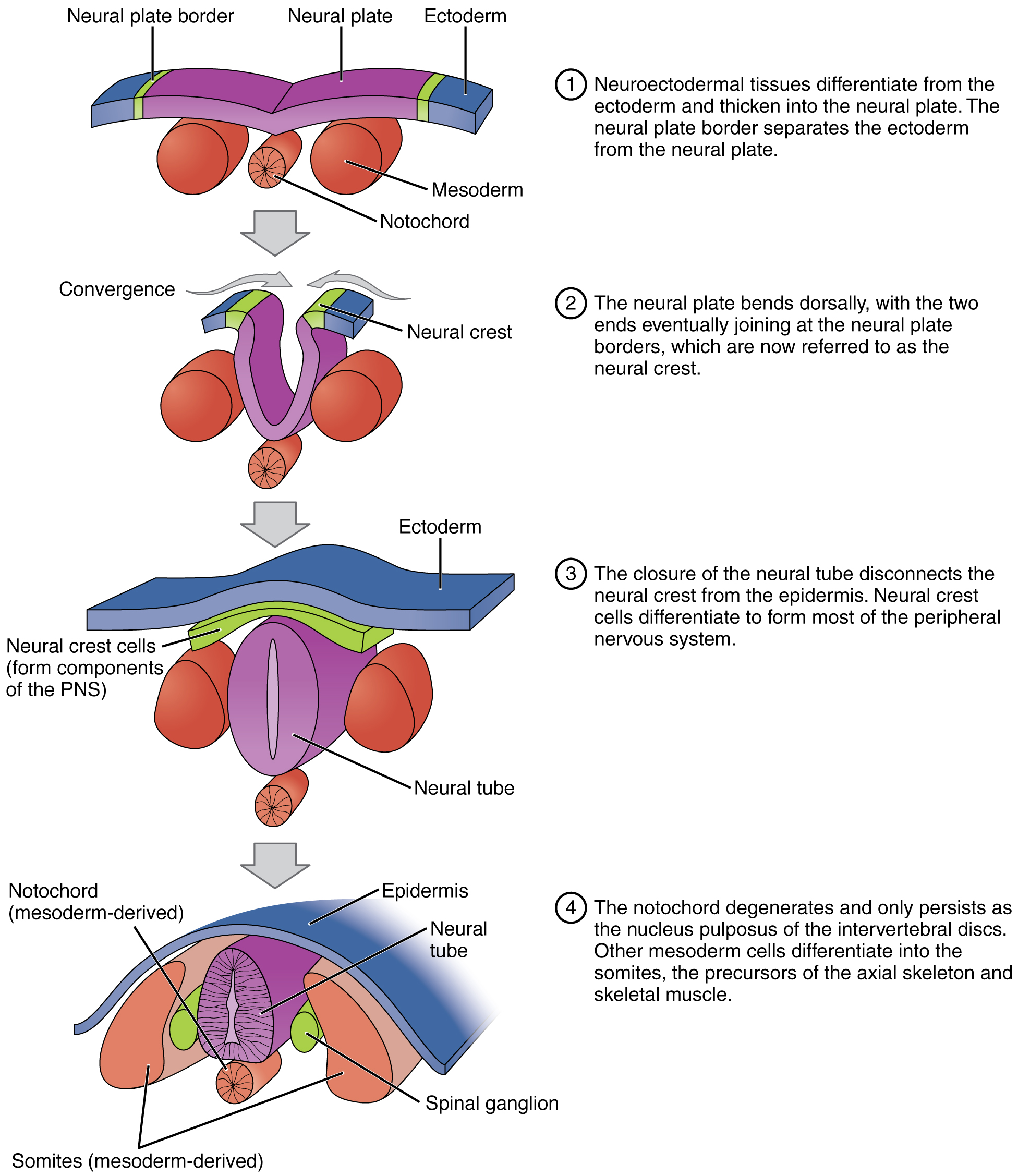
The mesoderm also differentiates into tissues that compose the cardiac muscle, smooth muscle, tissues within the kidneys, and red blood cells. The endoderm of vertebrates are the origins of other animal organs, including lungs, thyroid, and pancreas.
1.4 The Origin of Animals
The eukaryotes include a huge variety of single-celled (unicellular) organisms, mostly lumped together in the category protists, which includes any eukaryote that isn’t a plant, fungus, or animal. Animals are unique in that they are obligatory multicellular eukaryotes that obtain energy by ingesting food. Animals may have evolved only once from a unicellular ancestor: All known animals share so many molecular features that it seems certain that they have a single common origin.
Protists are neither plants, animals, nor fungi. They are predominantly unicellular microorganisms that may share certain morphological and physiological characteristics with animals or plants, or both.
Some protists have features of both animals and plants, such as the Euglena found in fresh and salt water. Most species of Euglena have chloroplasts with chlorophyll. Thus, when the euglena is exposed to sunlight it behaves as an autotroph. Conversely, in a food rich environment it behaves as a heterotroph ingesting particles of food by phagocytosis.
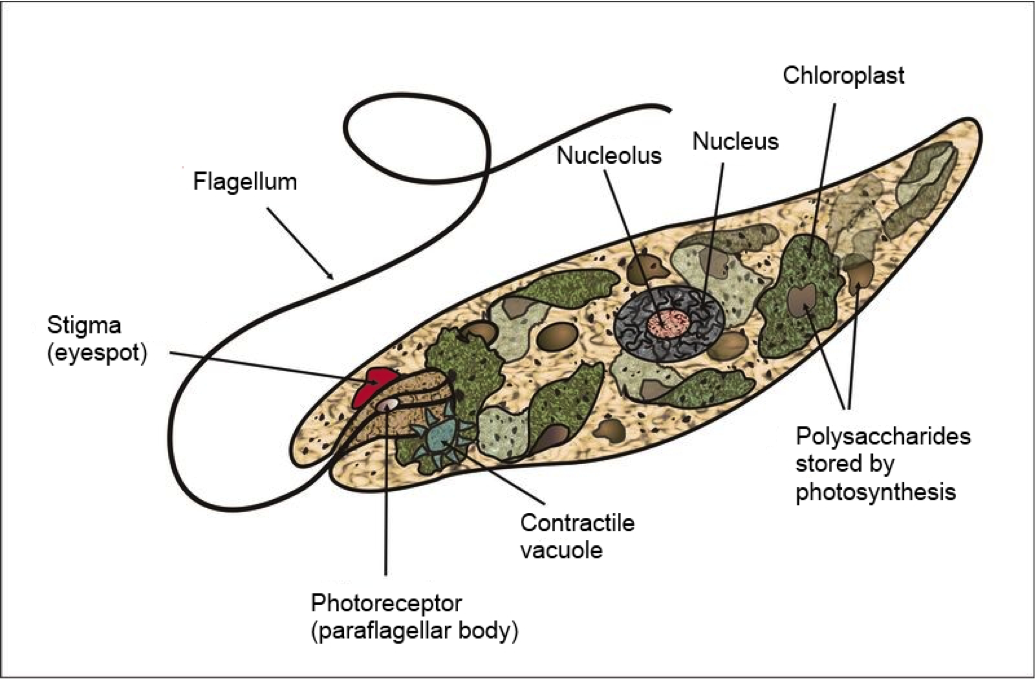
Many protists have animal-like characteristics: Many move, many are predators, and many have no cell wall. Protists with such animal-like characteristics are informally called protozoa. Examples of protozoa include the ciliate Paramecium, widespread in fresh water and ponds, pathogens such as Plasmodium, a flagellate parasite of blood that causes malaria, and Amoeba, a single celled organism that alters its shape by extending and retracting pseudopods (amoeboid movement). A relative of Amoeba, Entamoeba, causes the disease amoebic dysentery in humans and other animals.
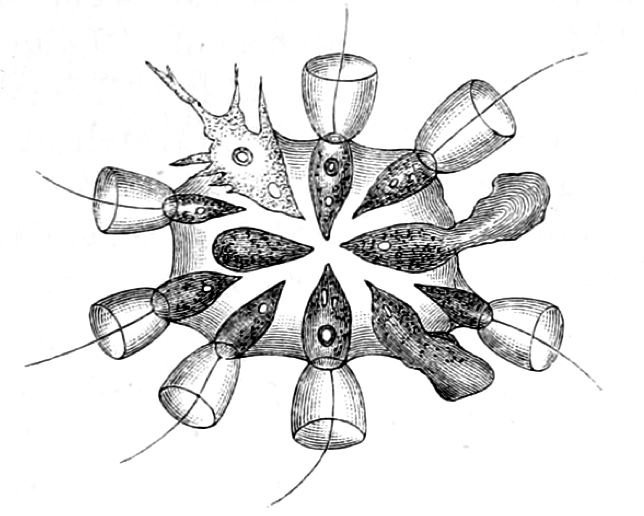
The closest living relatives of modern animals are the collared protists, the choanoflagellates, which live in water environments. This name comes from a flagellum surrounded by a collar of microvilli. There are approximately 140 species of choanoflagellates, some of which are single celled, while others form colonies attached to a solid substrate by a common stalk. They do not have chloroplasts and therefore are heterotrophs. The propelling movement of the flagellum creates water currents that trap bacteria and detritus against the collar of microvilli. The collar of microvilli acts as a net to catch the tiny prey, and the cell engulfs the prey by phagocytosis.

Genome sequencing studies on choanoflagellates revealed that they have many of the same cell signalling and adhesion molecules as animals. Thus, choanoflagellates are used as a model for reconstruction of the last unicellular ancestor of animals and are of particular interest to evolutionary biologists studying the origins of multicellularity in animals. Because choanoflagellates and metazoans are closely related, comparisons between the two groups promise to provide insights into the biology of their last common ancestor and the earliest events in animal evolution.
The Colonial Hypothesis. A fundamental question in the origin of animals is how unicellular organisms became multicellular, and how the cells of multicellular organisms further differentiated into specialized tissues. Presumably, the first multicellular organisms evolved from colonies of unicellular protists. Many modern flagellates live in colonies. According to this hypothesis, colonial cells became more and more interdependent by cell specialization to perform different functions. Specialized cells were interdependent as they could not survive on their own therefore the colony became a multicellular organism. As cells became more specialized, they differentiated further, forming tissues and then organs.
Most likely, animal cells originated in the salty fluid medium of the oceans. Therefore, it is quite possible that one of the first functional requirements was controlling intracellular fluid volume. By limiting the concentration of sodium ions, cells control their volume. The membrane of all animal cells contains an antiporter enzyme, the sodium-potassium adenosine triphosphatase (Na+/K+-ATPase) also known as the Na+/K+-pump that pumps three sodium ions out of the cell for every two potassium ions pumped into the cell. The Na+/K+-ATPase activity accounts for two important properties of animal cells:
- It maintains a low intracellular concentration of sodium, therefore it reduces the osmotic pressure controlling cell volume;
- It creates a difference in the amount of positively charged ions between the interior and the exterior side of the cell membrane, creating a charged environment termed membrane potential.
Osmosis is the passive diffusion of water into a compartment that has a greater concentration of solutes.
Sponges, phylum Porifera (Greek, “pore bearer”), are among the simplest animals known. They are sessile[5] with flagellated collar cells similar to the choanoflagellates, using water flow and filtering to capture their microscopic prey. Sponges have relatively few cell types and differentiated tissues; do not have digestive or circulatory systems, instead they rely on maintaining a constant water flow through their bodies to obtain food and oxygen and also to remove wastes (Figure 1.8). The shapes of their bodies often are asymmetric, adapted to maximize the efficiency of the water flow. Most species feed on bacteria and other food particles in the water but some species host photosynthetic microorganisms called endosymbionts: The sponge provides nitrogen, phosphorus, and carbon dioxide, while the endosymbionts provide energy in the form of fixed carbon and molecular oxygen. Other species of sponges that live in food-poor environments are carnivores and prey mainly on small crustaceans.

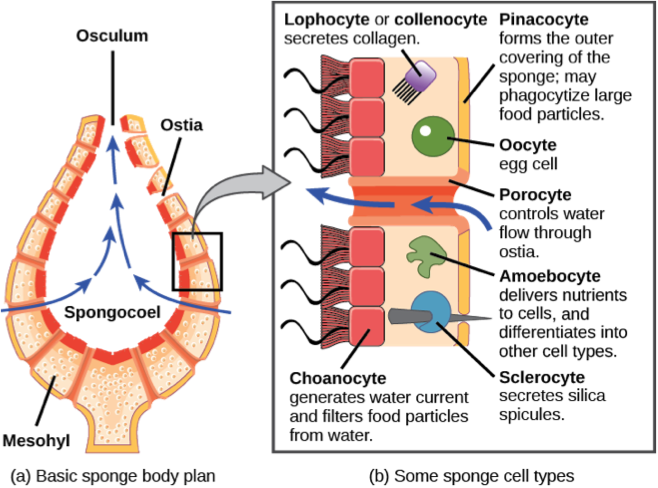
The body of the sponges consists of a type of connective tissue in the form of a gelatinous matrix, termed mesohyl, located between two thin layers of cells. Sponges have the capability of transforming some differentiated cells into other types of cells. This is a unique feature in the animal kingdom, since in eumetazoa, unspecialized cells differentiate into specialized cells. The collared cells of sponges, termed choanocytes, have the same basic structure as choanoflagellates. Collared cells are also found in other animal groups, such as ribbon worms, suggesting this was the morphology of the last common ancestor of all metazoa. Thus, it is considered quite possible that the common ancestor of all metazoan lineages in the Precambrian era was a sponge-like animal, and that differentiated tissues and embryonic development were innovations in the lineage that led to the eumetazoa after the split with the ancestor of the sponges.
Sponges do not have neurons or any other kind of nervous tissue. However, most species have pinacocytes, flat cells found on the outermost layer of the sponge, that are capable of contracting and expanding their water channels to create coordinated movement of the entire body. Some species of sponges can contract the osculum independently of the rest of the body. Cells of the osculum tissue coordinate their action through waves of calcium ions, similar to cell-to-cell communication in many tissues of complex animals. From an evolutionary perspective, the organization of multicellularity in sponges would have preceded the development of more specialized cells and the advanced mechanisms of interaction and communication characteristic of eumetazoa.
1.5 Gas Exchange and the Evolution of Animals
All animals must take up oxygen from the environment and distribute it to their cells and get rid of carbon dioxide, in a process called gas exchange. Animals obtain the energy needed for life by electron transfer from carbohydrates, fats and proteins to oxygen. Release of stored energy by electron transfer is termed cellular respiration. For more than one billion years, the earliest life forms had to use other electron acceptors for respiration, because the atmosphere contained only trace amounts of molecular oxygen (O2). Early ecological communities were simpler than current ones, presumably because a limitation of anaerobic respiration is that it cannot support high metabolic rates characteristic of complex forms of life such as animals.
Before oxygen came to make up a substantial part of air, all organisms were small. None of their parts was far from their external surface, facilitating the exchange of material from the environment with all parts of the body by simple diffusion.
Diffusion is the net movement of molecules or atoms from a region of high concentration to a region of low concentration.
A significant prerequisite for the first appearance of animals was the generation of atmospheric oxygen by cyanobacteria that use water as an electron donor for photosynthesis. The oxidation of water releases electrons and molecular oxygen (O2). The presence of molecular oxygen revolutionized the composition of life on earth: even though oxygen is a toxic molecule, the new ability to store and release a large amount of energy ultimately led to an explosion of biodiversity.
Small organisms are also highly vulnerable to larger predators. It is likely that a combination of changes in the physical environment, especially oxygen concentrations, and interactions between predators and their prey favoured increases in the size of many organisms. The steady increase in atmospheric concentrations of oxygen facilitated larger animals to evolve because they were able to obtain enough oxygen to maintain the metabolism of all of their cells.
Large unicellular organisms have lower surface-to-volume ratios. They can obtain oxygen by simple diffusion only if concentrations are higher that the concentrations that support small prokaryotic cells. Larger animals have evolved specialized structures, such gills and lungs to exchange gases, and circulatory systems to efficiently transport oxygen and carbon dioxide to and from their cells.
1.6 Cell Interaction and Tissues
The first hallmark of multicellularity was the formation a space between cells or interstitium (Latin, inter: between, and sistere: place) known as the interstitial space (IS). This space is filled with the interstitial fluid (IF) that bathes and surrounds the cell. In vertebrates and a couple of other animal groups, the circulatory system forms a separate compartment of vascular structures occupied by blood. Blood is composed of cells suspended in plasma fluid (see Figure 1.9). The IF and plasma are in interconnected compartments and both are in direct contact with cells, therefore, together are a wider fluid compartment termed the extracellular fluid (ECF).

In invertebrates, the IF and plasma combine to form the hemolymph, a blood-like fluid that fills the animal’s body interior and surrounds all cells. Overall, these fluid environments constitute the internal media, whose composition is tightly regulated.
The formation of extracellular space and the ECF protect cells from the harsh external environment to which the primitive animal cell colonies were exposed. Maintaining a complex animal body requires the ability to control the variables of the internal media, including nutrients, pH, temperature and concentration of electrolytes. Controlling the internal media facilitates the second hallmark in animal multicellularity: the differentiation of cells into specialized forms and the division of labor. Excitable cells, such as in muscle and neurons evolved further, developing into motor and sensory systems of visual, olfactory, auditory, and tactile perception.
The Extracellular Environment. The extracellular space is also occupied by a network of macromolecules forming the extracellular matrix (ECM). The ECM provides structural support for the tissue and gives the cells a place to attach. The ECM also performs biochemical reactions, as it includes enzymes, and additionally functions in communication between cells due to the presence of signalling molecules. The ECM can be a soft gel, a tough fibrous material, or even a rock-hard solid, depending on the specific tissue. A thin, sheet-like deposit of ECM is the basement membrane on which various kinds of sheets of cells – epithelia, mesothelia, and endothelia – rest.
The macromolecules that give the ECM its structure are typically glycosaminoglycans and glycoproteins produced by the neighbouring cells.
The protein collagen, which is a main component of the ECM, arose very early in metazoan evolution; It is produced by sponges and other primitive organisms and is the main structural protein in the interstitial space of a wide range of animals. In simple organisms the ECM forms the cuticle (outer skin) of roundworms and the shell of molluscs. In most complex organisms, collagen is mostly found in fibrous tissues such as tendons, ligaments, muscle and skin and is the most abundant protein in mammals (25% to 35% of the human body’s protein by weight).
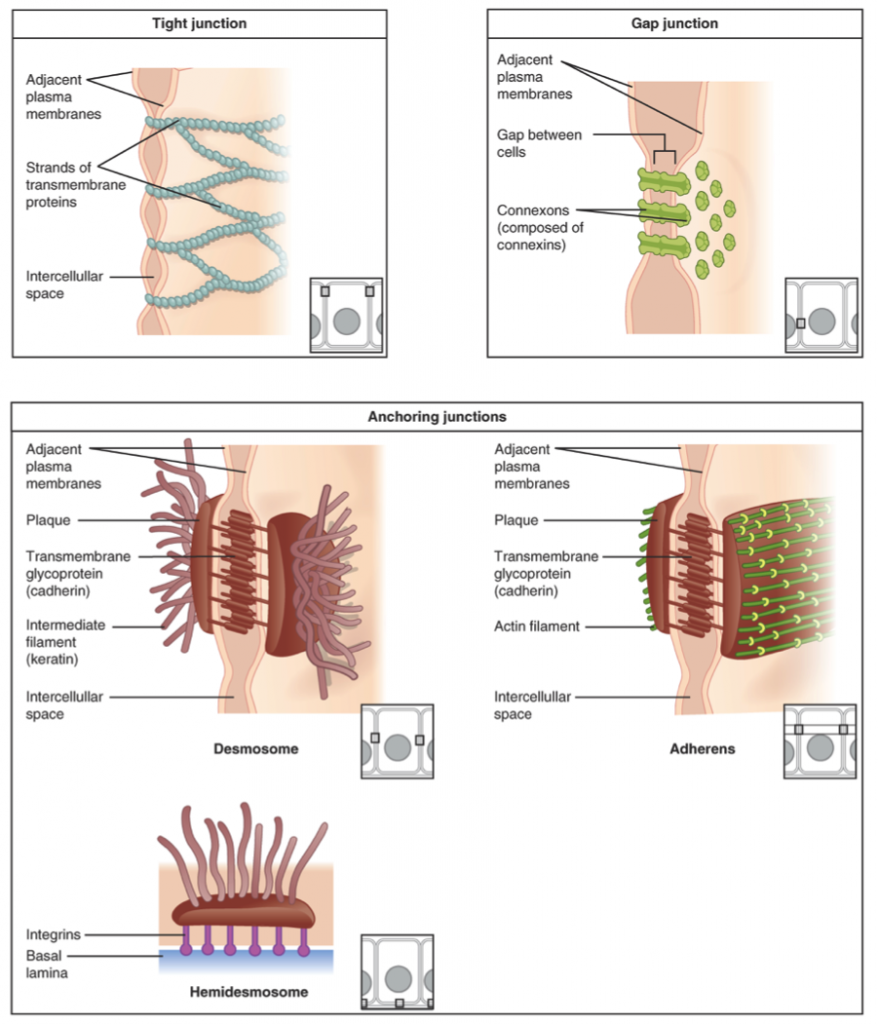
Cell Junctions. A crucial step in the transition to multicellularity was the development of structures that directly connect cells. Cells are connected through adhesion structures that are unique to animals. These structures, unique to animals, include the tight junctions, gap junctions, and anchoring junctions.
- Tight junctions or zona occludens – are rows of transmembrane proteins that bind to the corresponding rows of membrane proteins of the adjacent cell, forming a tight fence that separates the two sides of a sheet of cells. Thus, tight junctions form a virtually impermeable barrier to the flow of fluids, ions, and large molecules. Tight junctions are composed of two main groups of proteins, the claudins (Latin, claudere: to close) and the occludins. Intracellular membrane proteins anchor actin filaments of the cytoskeleton to claudins and occludins, mechanically joining the cytoskeletons of adjacent cells.
- Gap junctions or nexus junctions – are intercellular channels that directly communicate the cytoplasm of two cells, allowing passage of ions and small molecules. In vertebrates, the gap junctions are composed of connexin protein, while the unrelated but functionally similar innexins form the gap junctions of invertebrates.
- Anchoring junctions – Cells within tissues and organs are anchored to one another through a collection of molecules that link the cytoskeleton with the extracellular matrix. Three main types of anchoring junctions have been described in animal tissues: adherens junctions, desmosomes, and hemidesmosomes. They differ mainly in the cytoskeletal anchor protein as well as the transmembrane linker protein that extends through the cell membrane. Adherens junctions, also termed zonula adherens, intermediate junction, or belt desmosome are junctions whose cytoplasmic face is linked to the cytoskeleton filaments composed of the glycoprotein actin. They can form bands encircling the cell (zonula) or spots of attachment to the extracellular matrix (adhesion plaques). Desmosomes join cells through a group of proteins termed cadherins that join cells of the same tissue by homophilic binding attached to intermediate filaments of the cytoskeleton composed of keratin. A common proteome of identical cadherins termed desmogleins characterize cells of the same tissue allowing cells to recognize and bind each other. Hemidesmosomes are rivet-like structures on the inner basal surface of the keratinocytes the form the skin epidermis. Hemidesmosomes attach cells together through a group of proteins termed integrins connecting the basal face of a cell with a basal lamina of extracellular matrix.
1.7 Animal Tissues
Further levels of cellular organization provide multicellular organisms with distinct properties. The first level of organization was the integration of cells into tissues. A tissue is defined as group of cells that function together in a specialized and coordinated activity. Two or more tissues form an organ, which is defined as a structure that performs defined functions in the body. Different organs work together to perform a larger function, which defines a system.
The nervous/endocrine system is one example of an organ system. Another example is the cardiovascular, respiratory, and urinary system of organs. The spatial arrangement of organs into organ systems is part of the overall body plan, genetically determined and unique for each phylum of the animal kingdom. The body of a modern animal typically has over 200 different types of cells. Tissues are divided into four main categories with different functional specializations:
Muscle Tissue – This is composed of cells specialized to contract, generating force. Most animals have three kinds of muscle tissue:
- Skeletal muscle, or striated voluntary – attached to bone or exoskeletons, used for locomotion under voluntary control.
- Cardiac muscle, or striated involuntary – found only in the heart, under involuntary control.
- Smooth muscle – surrounds hollow tubes and cavities for propulsion of contents, under involuntary control.
Nervous Tissue – This is composed of cells termed neurons, specialized in generation and conduction of electrical signals in the animal body.
Epithelial Tissue – This is composed of densely packed sheets of cells that cover the body or organs and line the walls of body cavities. Epithelial cells are specialized in:
- Protection;
- Secretion and/or absorption of ions and organic molecules; and
- Secretion of chemical signalling molecules, for example hormones, growth factors, and neurotransmitters.
Epithelial cells are asymmetrical in that they are polarized[6], with different structures and proteins facing each of the two surfaces of the sheet of cells. They rest on basal lamina or basement membrane and can function as selective barriers. Epithelial cells can be found in the following main shapes:
- Cuboidal – similar width and height, like a cube
- Squamous – flattened
- Columnar – elongated
Arranged as:
- simple – one layer,
- stratified – multiple layers, or
- pseudostratified – one layer, with the nuclei staggered.

Connective Tissues – They are composed of cells, fibers and the ECM. They include blood, adipose, bone, cartilage, and loose and dense connective tissues. The ECM also forms the basement membranes that support cells. Basement membranes are sheet-like deposits of ECM over which epithelial cells rest. The ECM is also the main component of cartilage and may form calcified structures such as shells, bones, and spicules.
The ECM serves many other functions, including segregating tissues from one another and mediating intercellular communication. It regulates a cell’s dynamic behavior, and stores a wide range of cellular growth factors that can be rapidly liberated without de novo[7] synthesis. Formation of the ECM is an essential process for normal body growth and wound healing.
Many cell types comprise the ECM in different tissues, including fibroblasts, chondrocytes and osteoblasts, fibroblasts being the most common cell type in connective tissue. Fibroblasts produce glycosaminoglycans, proteoglycans and glycoproteins, the precursor components of the ECM. Chondrocytes, found in cartilages, produce the cartilaginous matrix. Osteoblasts are the cells that deposit bone.
1.8 Animal Body Plan
In biology, a body plan is defined as a collection of morphological features shared by a phylum[8]. Each species of multicellular organisms, from plants and fungus to the most advanced animal species, has a genetically determined body plan. Recognizably modern animal phyla first appear in marine fossils of the Cambrian explosion era about 550 million years ago, soon after the emergence of differentiated multicellular forms of animal life. Fossils from that time suggest that the way an animal’s body is laid out corresponds to a blueprint of body organization common to all animals in each phylum in existence today.
Symmetry and the “Pipe” Body Plan – Symmetry refers to the balanced distribution of duplicate body parts or shapes in the animal body. Simple animals such sponges are often asymmetrical, as they have irregular shapes that are adapted for maximal efficiency of water flow through the central cavity to trap food. The rest of the animal groups exhibit some form of symmetry, either radial or bilateral.
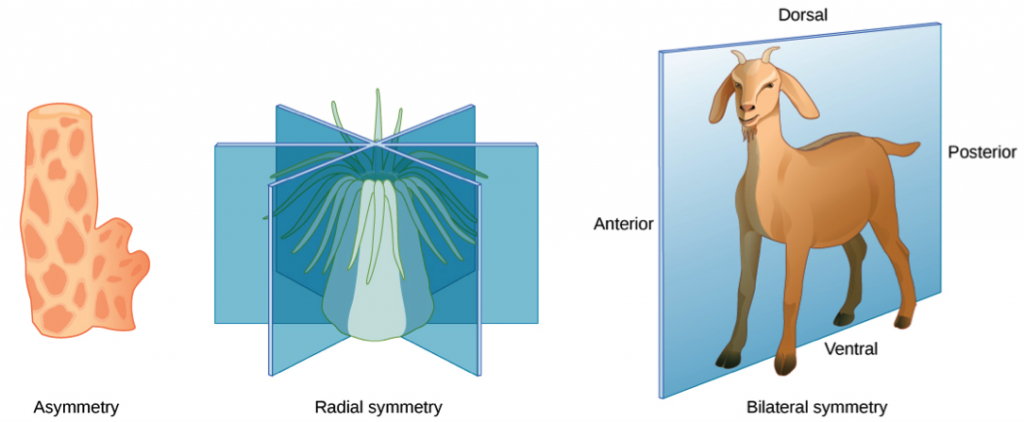
The radially symmetrical phyla Ctenophora (comb jellies) and Cnidaria (corals, sea anemones, and other jellies) are diploblastic animals. In radial symmetry, the body looks similar if rotated partway around a central axis. A radially symmetrical body can have top and bottom surfaces, but lack left and right sides, or front and back. The body of a jellyfish is shaped like a bell or umbrella, with a clear jellylike material filling most of the space between the upper and lower surfaces. Tentacles hang and wave from the bell margin. In the center of the undersurface, they have a mouth, from which they eat and discard waste.
The bodies of bilaterally symmetrical animals (mostly in the clade Bilateria) have mirror-image left and right sides. They have an anterior end with a specialized head with feeding and sensory organs, and typically a complex of digestive chambers and organs that connects with an anus or cloaca at the posterior end. This body configuration of an alimentary canal is similar to a tube, and is known as the pipe body plan.
This basic symmetry and organization gave ancient organisms an enormous advantage at survival and reproduction. All land animals descend from bilaterian ancestors with an alimentary canal allowing efficient nutrient capture, digestion, and waste disposal. Natural selection created an enormous range of variations on this basic configuration, for example with limbs, brains and skeletons – but the pipe model itself remains at its core. Thus, bilateral symmetry is common to diverse organisms from earthworms to humans.
An exception to the pipe plan of Bilateria are the echinoderms, a phylum of marine animals recognized by their radial symmetry, such as starfish, sea urchins, sand dollars and sea cucumbers. Echinoderms start life with bilateral symmetry, but become radially symmetrical by the time they become adults.
The evolution of bilateral symmetry and the formation of anterior and posterior ends led to the concentration an organized nervous system at the animal’s anterior end. This evolutionary trend is called cephalization (Greek: enkephalos, brain), while the increase in brain mass is called encephalization. From an evolutionary perspective, bilateral symmetry and the pipe body plan facilitated active mobility and increased sophistication of resource seeking, capture of prey and protection from predators, while radial symmetry remained for stationary and slow-moving species.
Body Segmentation – An animal’s shape, symmetry, number of body segments and limbs are all aspects of the genetically determined body plan. The body of all members of the three large animal phyla Annelida, Arthropoda, and Chordata have segmented body plans, meaning that they are constructed of a linear series of repeating parts or segments, each being formed in sequence along the cranial-caudal (head to tail) axis of the embryo.
Advantages of body segmentation include free movement, development and regeneration of different body parts.
The Body Plan of Arthropods (Phylum Arthrophoda, from Greek, arthron, “joint”, and podos “foot” or “leg”). These are invertebrates with an exoskeleton, a segmented body and jointed appendages. Members of this group include the insects, arachnids, and crustaceans. They have jointed limbs and cuticles of α-chitin. The cuticles of crustaceans are also mineralized with calcium carbonate. The rigid cuticle inhibits growth, so it is periodically replaced by moulting.
Body Cavities – These are the fluid-filled spaces that separate the internal organs from the outer body. Most animals have body cavities collectively termed the coelom, a definition that excludes blood and lymph vessels. The coelom is lined by mesodermal tissue. The coelom facilitates internal organ movement, protecting them from injury.
- Coelomates – Animals with a well-developed coelom are known as coelomates. The coelom is formed by a fluid-filled cavity lined by mesoderm-origin tissue.
- Pseudocoelomates – Animals with a fluid-filled cavity not completely lined by tissue derived from mesoderm are called pseudocoelomates. This group includes rotifers and roundworms (nematodes).
- Acoelomates – Several different kinds of animals with very simple body plans have no body cavities, for which they are known as acoelomates. Instead of fluids, acoelomates contain a connective tissue of spongy texture that fills the body.
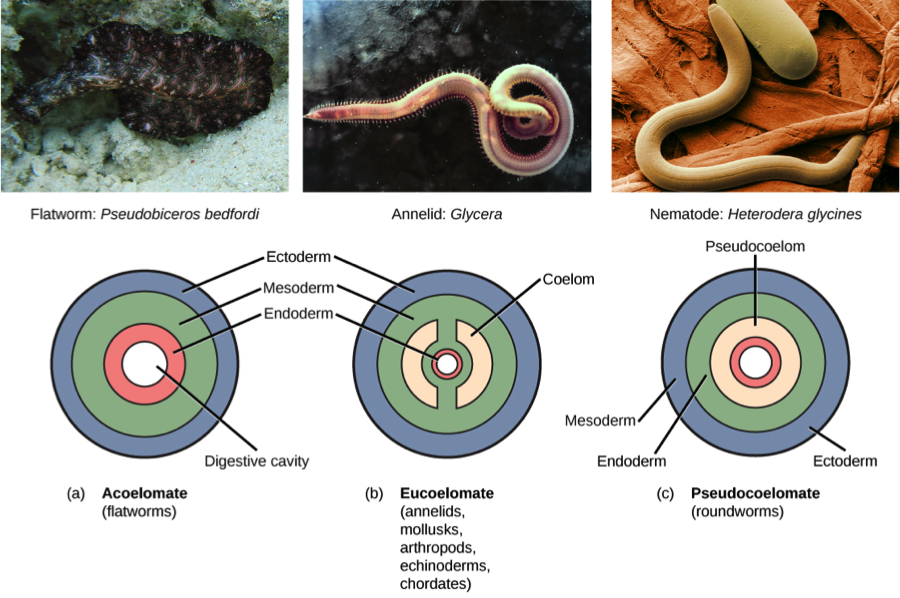
The development of body cavities provided several evolutionary advantages to animal species, including independent movement of body wall and enclosed organs, and more space for the development of complex organs and organ systems. The coelom fluid protects internal organs and provide nutrients to cells and serves for transport of gases and waste products.
Among coelomate animals, the coelom is typically the most obvious in the embryo because the coelom is modified in various ways during development into an adult:
- In earthworms and some other soft-bodied invertebrates, the coelom is a fluid-filled cavity in the form of a hydrostatic skeleton, which is surrounded by muscles that gives support and shape to the body and function as a locomotive apparatus.
- In arthropods and most molluscs the coelom is reduced to the pericardial cavity, that displaces the coelom to form an open circulatory system termed the hemocoel, filled with the hemolymph that contacts the tissue directly.
- In vertebrates, the mesoderm lines the coelom with thin tissue termed peritoneum. The inner and outer layers of the peritoneum fold and attach the stomach, small intestine, pancreas, spleen and other organs to the posterior wall of the abdomen.
Simple diploblastic animals such as sponges, cnidarians and ctenophores are acoelomates. Acoelomates also include some triploblastic species such as flatworms, bilaterally symmetrical animals that have more complex internal organs than cnidarians and ctenophores. Flatworms have no body cavity therefore lack organs for excreting metabolic wastes or for transporting oxygen to internal tissues. The flat body shape exposes all their cells close to water in the surrounding environment or in their gut. They have a relatively simple excretory apparatus that functions mainly to maintain osmotic balance with their surroundings. Flatworms exchange gases and eliminate nitrogenous waste by diffusion across the body surface. Thus, evolutionary progression of flatworms was limited by their ability to absorb nutrients, dispose of waste, and exchange gases.
1.9 The Chordates
The chordates are a phylum of Bilateria characterized by a having a notochord at some point of their life cycle. Some chordates have a notochord as adults, while many have the notochord only in the embryo stage. The notochord is a semi-rigid structure of cartilage along the inside of the body. The notochord functions as an attachment point for muscles, such that the body bends without shortening when the muscles contract. Chordates also have a hollow dorsal nerve cord. The dorsal nerve cord functions as the main communication stem of the nervous system.
The phylum Chordata includes the subphylum Vertebrata (Latin, vertebratus: backboned) that comprises all animals with an internal skeleton. In most vertebrates, the notochord is replaced by bone. During development, the notochord is replaced by a hollow structure of interlocking vertebrae that provides support and contains the nerve cord. Bone is a mineralized connective tissue that, in addition to supporting the body, performs a variety of other functions. Bone is a metabolically active tissue that contains living cells that grow with the animal. Skeletal cells are involved in mineralization of the tissue and secrete hormones and growth factors. Bones also contains the bone marrow a specialized connective tissue that produces the red blood cells (erythrocytes), a process termed the erythropoiesis (Greek, erythro: red and poiesis: to make).
In the chondrichthyes (Greek: chondr, cartilage and ichthys: fish) class of fish, that include the shark, the notochord present in young is gradually replaced by cartilage rather than bone. They do not have bone marrow. Blood cells are produce in the spleen and in a tissue that located around the gonads, termed the epigonal organ.
Vertebrates are coelomates that contain diverse internal organs located in two cavities, the thoracic and the abdominal. They include a heart with at least two chambers, a liver, kidneys and endocrine glands. The liver is exclusive to vertebrates, while the rest of the internal organs are more complex than analogous structures in other groups of animals. Basal vertebrates breathe with gills located right behind the head. In amphibians and some bony fishes, the larvae bear external gills, branching off from the gill arches. During adulthood in amphibians, external gills are reduced and their functions are taken over by lungs.
Most land vertebrates have two pairs of limbs[9]. Through evolution, some vertebrates developed innovations of these appendages adapted to their specific environment: water, land or air. Fins are thin appendages attached the body; typically function as foils to generate thrust, steer or stabilize motion while traveling in a fluid media such as water. Fins also increase surface areas for heat exchange. Amphibians were the first creatures to evolve limbs suitable for life on land. Most animals use limbs for locomotion, such as walking, running, or climbing. Some animals can use their front limbs (or upper limbs in humans) to carry and manipulate objects. Human legs and feet are specialized for two-legged locomotion, while most other mammals walk and run on all four limbs. Birds developed the front limbs into wings that enabled them to fly. Birds are covered with feathers, which are epidermal growths that form the distinctive outer covering, or plumage. Feathers aid in flight, thermal insulation, and waterproofing. The colouration of feathers helps in communication and camouflage.
The mammals (Latin, mamma: breast), a group of animals that evolved from four-legged ancestors, characterized by a mammary gland, a female organ that produces milk to feed young offspring. Mammals are endothermic, meaning that they usually maintain a constant body temperature much warmer than their environment. Mammals are also amniotes, meaning a membrane, the amnion[10] (Greek, amnion), closely covers the embryo. Most mammals retain the fertilized egg within the mother, but a couple of species lay their eggs on land. The largest groups of mammals are the placentals, which nourish the fetus before birth in the mother’s uterus through a specialized embryonic organ attached to the uterus wall, the placenta. Mammals are distinguished from reptiles and birds by the development of a new six-layered region of the brain’s cortex (Latin, cortex: rind) called the neocortex (Greek, neo: new). Three quarters of the human brain’s mass is neocortex. Mammals are also characterized by the presence of skin hair and three middle ear bones in the auditory system. The mammals include the largest surviving vertebrates.
1.10 Summary
- The word “animal” refers to any member of the kingdom animalia, also known as metazoa;
- All animals are multicellular as adults;
- Most animals are heterotrophs (use food as a source of energy stored in organic carbon compounds);
- Most animals move and respond to stimuli;
- Most animal species consist of male and female individuals that reproduce sexually by fusing a sperm cell with an egg cell;
- The cell formed by fusion of sperm and egg divides, resulting in cells that differentiate to form tissues and organs;
- Cells are organized into tissues, and tissues are organized into organs;
- Tissues are divided into four main categories: muscle, nervous, connective, and epithelial tissues;
- The interstitial fluid between cells is salty, suggesting that animals evolved from colonies of unicellular protists in the sea;
- The cell membrane of all animal cells contains an antiporter enzyme, the sodium-potassium adenosine triphosphatase, that pumps three sodium ions out of the cell for every two potassium ions into the cell (actually, if the Na+/K+-ATPase is pumping against a concentration gradient, the number of ions transported per ATP molecule hydrolysed can be less);
- The extracellular space is also occupied by macromolecules, such as glycosaminoglycans and collagen that form the extracellular matrix;
- Cell adhesions are crucial to multicellularity, and include tight junctions, gap junctions, and desmosomes;
- In most animals, one side of the body is almost a mirror image of the other side. This is called bilateral symmetry;
- Many bilaterians have body cavities collectively termed the coelom;
- One group of coelomates, the chordates, have a semi rigid structure of cartilage, the notochord, in the body of the embryo, along with a hollow dorsal nerve cord;
- The vertebrates are a group of chordates where the notochord is replaced by interlocking vertebrae;
- The amniotes are a group of vertebrates where a membrane, the amnion, closely covers the embryo;
- The largest vertebrates are the mammals, characterized by having mammary glands;
- Mammals are endothermic, meaning that they usually maintain a constant body temperature much warmer than their environment;
- The largest group of mammals are the placentals, which nourish the fetus before birth in the mother’s uterus through the placenta, a specialized embryonic organ attached to the uterus wall;
1.11 Suggested Readings
Biology, OpenStax College. 21 June 2012. http://cnx.org/content/col11448/latest/
Biology – Exploring the Diversity of Life (3rd Canadian Edition) by Russell et al., Nelson Education Ltd., 2016.
1.12 Glossary
CAM – cell adhesion molecules
ECF – extracellular fluid
ECM – extracellular matrix
IF – interstitial fluid
IS – interstitial space
Na+/K+ ATPase – sodium-potassium adenosine triphosphatase
- “And the Lord God formed man of the dust of the ground, and breathed into his nostrils the breath of life; and man became a living soul.” Genesis 2:7. ↵
- “Hermaphrodite”, from the Latin hermaphroditus, derived from the ancient Greek word hermaphroditos, a character described in Greek mythology as the son of Hermes and Aphrodite who becomes joined in one body with a nymph while bathing. ↵
- There are a few exceptions, such as the giant clam Tridachna gigas and the green sea slug Elysia chlorotica, and, most famously, corals. The bodies of these animals contain symbiotic algae that carry out photosynthesis, providing the animals with a supplemental source of carbon or energy. All of these animals eat as well – unlike plants, they aren’t pure autotrophs. ↵
- Symbiosis refers to the interaction between two different biological species. It is generally accepted that symbiotic relationship is mutually beneficial. ↵
- Sessile” means “immobile”, attached in place to a solid substrate for a long time, or even permanently. A condition of some aquatic animals, such as barnacles and corals. ↵
- Refer to two opposite poles of the epithelial cells, one pole facing the basement membrane, while the other pole faces a lumen or cavity. ↵
- Latin term used to indicate synthesis of a cell product from simple molecules such as sugars or amino acids ↵
- Phylum (plural: phyla) is a taxonomic rank below kingdom and above class. ↵
- Some vertebrates, including terrestrial and aquatic forms, have only one pair of appendages or even no visible appendages, such as fins, legs, or arms, for which they are termed tetrapods (Greek, tetra: four; podēs: feet, or four-footed). ↵
- Reptiles and birds are amniotes too. ↵
